Citation: Yufei Wang, Huidong Cheng, Qingjian Li. Dynamic analysis of wild and sterile mosquito release model with Poincaré map[J]. Mathematical Biosciences and Engineering, 2019, 16(6): 7688-7706. doi: 10.3934/mbe.2019385

| [1] | K. Ciesielski, On stability in impulsive dynamical systems, B. Pol. Acad. Sci. Math., 52 (2010), 81–91. |
| [2] | Z. Shi, Y. Li and H. Cheng, Dynamic analysis of a pest management smith model with impulsive state feedback control and continuous delay, Mathematics , 7 (2019), 591. |
| [3] | T. Zhang, X. Liu, X. Meng, et al., Spatio-temporal dynamics near the steady state of a planktonic system, Comput. Math. Appl., 75 (2018), 4490–4504. |
| [4] | Y. Li, H. Cheng, J. Wang, et al., Dynamic analysis of unilateral diffusion Gompertz model with impulsive control strategy, Adv. Differ. Equ., 2018 (2018), 32. |
| [5] | J. Wang, H. Cheng, H. Liu, et al., Periodic solution and control optimization of a prey-predator model with two types of harvesting, Adv. Differ. Equ., 2018 (2018), 1687–1847. |
| [6] | N. Gao, Y. Song, X. Wang, et al., Dynamics of a stochastic SIS epidemic model with nonlinear incidence rates, Adv. Differ. Equ., 2019 (2019), 41. |
| [7] | Y. Li, H. Cheng and Y. Wang, A Lycaon pictus impulsive state feedback control model with Allee effect and continuous time delay, Adv. Differ. Equ., 2018 (2018), 1687–1847. |
| [8] | L. Zhang, Y. Wang, M. Khalique, et al., Peakon and cuspon solutions of a generalized Camassa-Holm-Novikov equation, J. Appl. Anal. Comput., 8 (2018). |
| [9] | Z. Shi, J. Wang, Q. Li, et al., Control optimization and homoclinic bifurcation of a preyCpredator model with ratio-dependent, Adv. Differ. Equ., 7 (2019), 591. |
| [10] | K. Liu, T. Zhang and L. Chen, State-dependent pulse caccination and therapeutic strategy SI epi-demic model with nonlinear incidence rate, Comput. Math. Method. M., 2019 (2019), 10. |
| [11] | W. Lv and F. Wang, Adaptive tracking control for a class of uncertain nonlinear systems with infinite number of actuator failures using neural networks, Adv. Differ. Equ., 2017 (2017), 374. |
| [12] | H. Liu and H. Cheng, Dynamic analysis of a prey-predator model with state-dependent control strategy and square root response function, Adv. Differ. Equ., 2018 (2018), 63. |
| [13] | F. Wang, B. Chen, Y. Sun, et al., Finite time control of switched stochastic nonlinear systems, Fuzzy Set. Syst., 365 (2019), 140–152. |
| [14] | X. Meng, F. Li and S. Gao, Global analysis and numerical simulations of a novel stochastic eco-epidemiological model with time delay, Appl. Math. Comput., 339 (2018), 701–726. |
| [15] | Y. Ren, M. Tao, H. Dong, et al., Analytical research of ( 3 + 1 )-dimensional Rossby waves with dissipation effect in cylindrical coordinate based on Lie symmetry approach, Adv. Differ. Equ., 9 (2019), 13. |
| [16] | J. Wang, H. Cheng, X. Meng, et al., Geometrical analysis and control optimization of a predator-prey model with multi state-dependent impulse, Adv. Differ. Equ., 2017 (2017), 252. |
| [17] | Y. Li, Y. Li, Y. Liu, et al., Stability analysis and control optimization of a prey-predator model with linear feedback control, Discrete Dyn. Nat. Soc., 2018 (2018), 12. |
| [18] | G. Liu, X. Wang and X. Meng, Extinction and persistence in mean of a novel delay impulsive stochastic infected predator-prey system with jumps, Complexity, 2017 (2017), 115. |
| [19] | F. Wang and X. Zhang, Adaptive finite time control of nonlinear systems under time-varying actu-ator failures, IEEE T. Syst., (2019). |
| [20] | J. Li, L. Cai and Y. Li, Stage-structured wild and sterile mosquito population models and their dynamics, J. Biol. Dynam., 11 (2016), 1. |
| [21] | W.Mylene, L.Georges, M.Knud, etal., Comparativegenomics: Insecticideresistanceinmosquito vectors, Nature, 423 (2003), 136–7. |
| [22] | L. Zhang and M. Khalique, Classification and bifurcation of a class of second-order ODEs and its application to nonlinear PDEs, Discrete Cont. Dyn. S, 11 (2017), 777–790. |
| [23] | F, Liu, Continuity and approximate differentiability of multisublinear fractional maximal func-tions, Math. Inequal. Appl., 21 (2018), 25–40. |
| [24] | P. Stiling, Biological control by natural enemies, Ecology, 73 (1992), 1520–1520. |
| [25] | A. C. Redfield, The biological control of chemical factors in the environment, Sci Prog, 11 (1960), 150–170. |
| [26] | Y. Li and X. Liu, H-index for nonlinear stochastic systems with state and input dependent noises, Int. J. Fuzzy Syst., 20 (2018), 759–768. |
| [27] | F. Zhu, X. Meng and T. Zhang, Optimal harvesting of a competitive n-species stochastic model with delayed diffusions, Math. Biosci. Eng., 16 (2019), 1554–1574. |
| [28] | L. Marshall, J. W. Miles and A. Press, Integrated mosquito control methodologies, (1983). |
| [29] | H. J. Barclay, Models for the sterile insect release method with the concurrent release of pesticides, Ecol. Model., 11 (1980), 167–177. |
| [30] | Y. Li and X. Meng, Dynamics of an impulsive stochastic nonautonomous chemostat model with two different growth rates in a polluted environment, Discrete Dyn. Nat. Soc., 2019 (2019), 15. |
| [31] | Y. Liu, H. Dong and Y. Zhang, Solutions of a discrete integrable hierarchy by straightening out of its continuous and discrete constrained flows, Anal. Math. Phys., 2 (2018), 1–17. |
| [32] | W. Klassen, C. F. Curtis, W. Klassen, et al., Sterile Insect Technique, (1989). |
| [33] | M. Q. Benedict and A. S. Robinson, The first releases of transgenic mosquitoes: an argument for the sterile insect technique, Trends Parasitol., 19 (2003), 349–355. |
| [34] | F. Liu, Q. Xue and K. Yabuta, Rough maximal singular integral and maximal operators supported by subvarieties on Triebel-Lizorkin spaces, Nonlinear Anal., 171 (2018), 41–72. |
| [35] | T. Feng, Z. Qiu, X Meng, et al., Analysis of a stochastic HIV-1 infection model with degenerate diffusion, Appl. Math. Comput., 348 (2019), 437–455. |
| [36] | L. Alphey, M. Benedict, R. Bellini, et al., Sterile-insect methods for control of mosquito-borne diseases: an analysis, Vector Borne Zoonot. Dis, 10 (2010), 295–311. |
| [37] | Y. Li, W. Zhang and X.Liu, H-index for discrete-time stochastic systems with Markovian jump and multiplicative noise, Automatica, 90 (2018), 286–293. |
| [38] | L. Zhang, Y. Wang, M. Khalique, et al., Peakon and cuspon solutions of a generalized Camassa-Holm-Novikov equation, J. Appl. Anal. Comput., 8 (2018), 1938–1958. |
| [39] | J. Li and Z. Yuan, Modelling releases of sterile mosquitoes with different strategies, J. Biol. Dyn., 9 (2015), 1–14. |
| [40] | H. Barclay and M. Mackauer, The sterile insect release method for pest control: a density-dependent model, Environ. Entomol., 9 (1980), 810–817. |
| [41] | F. Wang, B. Chen, Y. Sun, et al., Finite time control of switched stochastic nonlinear systems, Fuzzy Set. Syst., (2018). |
| [42] | H. Qi, X. Leng, X. Meng, et al., Periodic Solution and Ergodic Stationary Distribution of SEIS Dynamical Systems with Active and Latent Patients, Qual. Theory Dyn. Syst., 18 (2019). |
| [43] | M. Huang, X. Song and J. Li, Modelling and analysis of impulsive releases of sterile mosquitoes, J. Biol. Dynam., 11 (2017), 1147. |
| [44] | J. C. Panetta, A mathematical model of periodically pulsed chemotherapy: Tumor recurrence and metastasis in a competitive environment, B. Math. Biol., 58 (1996), 425–447. |
| [45] | S. Tang, B. Tang, A. Wang, et al., Holling II predator–prey impulsive semi-dynamic model with complex Poincaré map, Nonlinear Dynam., 81 (2015), 1575–1596. |
| [46] | D. R. J. Chillingworth, An Introduction to Chaotic Dynamical Systems by Robert L. Devaney, 1986. |
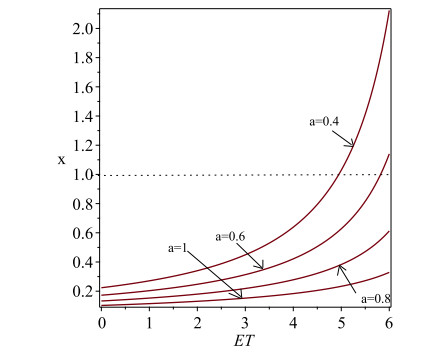
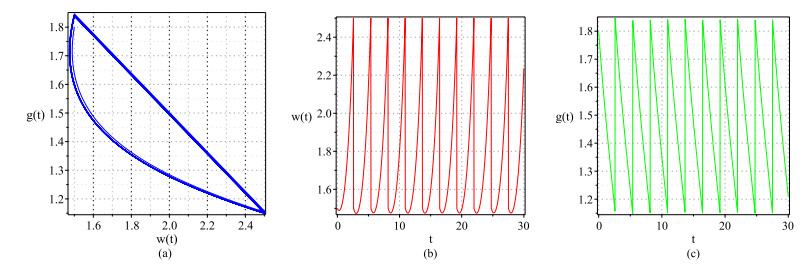
Figures(6)

Yufei Wang, Huidong Cheng, Qingjian Li. Dynamic analysis of wild and sterile mosquito release model with Poincaré map[J]. Mathematical Biosciences and Engineering, 2019, 16(6): 7688-7706. doi: 10.3934/mbe.2019385







 DownLoad:
DownLoad: