Citation: Shuyang Xue, Meili Li, Junling Ma, Jia Li. Sex-structured wild and sterile mosquito population models with different release strategies[J]. Mathematical Biosciences and Engineering, 2019, 16(3): 1313-1333. doi: 10.3934/mbe.2019064

| [1] | W. C. Allee, The Social Life of Animals, 2nd edition, Beacon Press, Boston, 1958. |
| [2] | L. Alphey, M. Benedict, R. Bellini, et al., Sterile-insect methods for control of mosquito-borne diseases: an analysis, Vector-Borne Zoonotic Dis., 10 (2010), 295–311. |
| [3] | H. J. Barclay, The sterile insect release method on species with two-stage life cycles, Res. Popul. Ecol., 21 (1980), 165–180. |
| [4] | H. J. Barclay, Pest population stability under sterile releases, Res. Popul. Ecol., 24 (1982), 405– 416. |
| [5] | H. J. Barclay, Modeling incomplete sterility in a sterile release program: Interactions with other factors, Popul. Ecol., 43 (2001), 197–206. |
| [6] | H. J. Barclay, Mathematical models for the use of sterile insects, In Sterile Insect Technique. Principles and Practice in Area-Wide Integrated Pest Management (eds. V.A. Dyck, J. Hendrichs, and A. S. Robinson), Springer, Heidelberg, 2005, 147–174. |
| [7] | H. J. Barclay and M. Mackauer, The sterile insect release method for pest control: A density dependent model, Environ. Entomol., 9 (1980), 810–817. |
| [8] | L. Cai, S. Ai and G. Fan, Dynamics of delayed mosquitoes populations models with two different strategies of releasing sterile mosquitoes, Math. Bios. Eng., 15 (2018), 1181–1202. |
| [9] | L. Cai, S. Ai and J. Li, Dynamics of mosquitoes populations with different strategies for releasing sterile mosquitoes. SIAM J. Appl. Math., 74 (2014), 1786–1809. |
| [10] | J. D. Charlwood and M. D. R. Jones, Mating behaviour in the mosquito, Anopheles gambiae s.l. I. Close range and contact behaviour, Phys. Entomol., 4 (1979), 111–120. |
| [11] | A. N. Davis, J. B. Gahan, D. E. Weidhaas, et al., Exploratory studies on gamma radiation for the sterilization and control of Anopheles quadrimaculatus, J. Econ. Entomol., 52 (1959), 868–870. |
| [12] | B. Dennis, Allee effects: population growth, critical density, and the chance of extinction, Nat. Res. Model., 3 (1989), 481–538. |
| [13] | Y. Dumont and J. M. Tchuenche, Mathematical studies on the sterile insect technique for the Chikungunya disease and Aedes albopictus, J. Math. Biol., 65 (2012), 809–854. |
| [14] | C. Dye, Models for the population dynamics of the yellow fever mosquito, Aedes aegypti, J. Anim. Ecol., 53 (1984), 247–268. |
| [15] | L. Esteva and H. M. Yang, Mathematical model to assess the control of Aedes aegypti mosquitoes by the sterile insect technique, Math. Bios., 198 (2005), 132–147. |
| [16] | J. Farkas and S. Gourley, Modelling Wolbachia infection in a sex-structured mosquito population carrying West Nile virus, J. Math. Biol., 75 (2017), 621–647. |
| [17] | J. C. Flores, A mathematical model for wild and sterile species in competition: Immigration, Phys. A, 328 (2003), 214–224. |
| [18] | L. Gomulski, Polyandry in nulliparous Anopheles gambiae mosquitoes (Diptera: Culicidae), Bull. Entomol. Res., 80 (1990), 393–396. |
| [19] | M. Huang, J. Lou, L. Hu, et al., Assessing the e ciency of Wolbachia driven Aedes mosquito suppression by delay differential equations, J. Theor. Biol., 440 (2018), 1–11. |
| [20] | E. F. Knipling, Possibilities of insect control or eradication through the use of sexually sterile males, J. Econ. Entomol., 48 (1955), 459. |
| [21] | E. F. Knipling, The basic principles of insect population suppression and management, Agriculture Handbook, US Department of Agriculture, Washington, DC, 512 (1979). |
| [22] | E. F. Knipling, Sterile insect technique as a screwworm control measure: the concept and its development, In Symposium on Eradication of the Screwworm from the United States and Mexico (ed. O.H. Graham), Misc. Pub. Entomol. Soc. Am., College Park, MD, 62 (1985), 4–7. |
| [23] | G. C. LaBrecque, D. W. Meifert and C. N. Smith, Mating competitiveness of chemosterilized and normal male house flies, Science, 136 (1962), 388–389. |
| [24] | J. Li, Malaria model with stage-structured mosquitoes, Math. Biol. Eng., 8 (2011), 753–768. |
| [25] | J. Li, Modeling of transgenic mosquitoes and impact on malaria transmission, J. Biol. Dyna., 5 (2011), 474–494. |
| [26] | J. Li, New revised simple models for interactive wild and sterile mosquito populations and their dynamics, J. Biol. Dyna., 11 (2017), 316–333. |
| [27] | J. Li, L. Cai and Y. Li, Stage-structured wild and sterile mosquito population models and their dynamics, J. Biol. Dyna., 11 (2017), 79–101. |
| [28] | J. Li and Z. Yuan, Modeling releases of sterile mosquitoes with different strategies, J. Biol. Dyna., 9 (2015), 1–14. |
| [29] | Y. Li and J. Li, Discrete-time models for releases of sterile mosquitoes with Beverton-Holt-type of survivability, Ricerche mat., 67 (2018), 141–162. |
| [30] | J. Lu and J. Li, Dynamics of stage-structured discrete mosquito population, J. Appl. Anal. Comput., 1 (2011), 53–67. |
| [31] | A. Mishra, B. Ambrosio and S. Gakkhar, A network model for control of Dengue epidemic using sterile insect technique, Math. Bios. Eng., 4 (2018), 441–460. |
| [32] | J. D. Mumford, Application of Benefit/Cost Analysis to Insect Pest Control Using the Sterile Insect Technique, In Sterile Insect Technique. Principles and Practice in Area-Wide Integrated Pest Management (eds. V.A. Dyck, J. Hendrichs, and A. S. Robinson), Springer, Heidelberg, 2005, 481–498. |
| [33] | S. J. Schreiber, Allee effect, extinctions, and chaotic transients in simple population models, Theor. Popul. Biol., 64 (2003), 201–209. |
| [34] | M. W. Steven, R. Pejman and M. S. Steven, Modelling pulsed releases for sterile insect technique: fitness costs of sterile and transgenic males and the effects on mosquito dynamics, J. Appl. Eco., 47 (2010), 1329–1339. |
| [35] | W. Takken, C. Costantini, G. Dolo, et al., Mosquito mating behaviour, In Bridging Laboratory and Field Research for Genetic Control of Disease Vectors (eds, B.G.J. Knols and C. Louis), Springer, Heidelberg, 2006, 183–188. |
| [36] | R. C. A. Thome, H. M. Yang and L. Esteva, Optimal control of Aedes aegypti mosquitoes by the sterile insect technique and insecticide, Math. Bios., 223 (2010), 12–23. |
| [37] | F. Tripet. Y. T. Toure, G. Dolo, et al., Frequency of multiple inseminations in field-collected Anopheles gambiae females revealed by DNA analysis of transferred sperm, Am. J. Trop. Med. Hyg., 68 (2003), 1–5. |
| [38] | WHO, Malaria, 2018, Available from: http://www.who.int/news-room/factsheets/ detail/malaria. |
| [39] | Wikipedia, Sterile Insect Technique, 2018, Available from: http://en.wikipedia.org/wiki/ Sterile/insect/technique. |
| [40] | J. Yu, Modelling mosquito population suppression based on delay differential equations SIAM J. Appl. Math., 78 (2018), 3168–3187. |
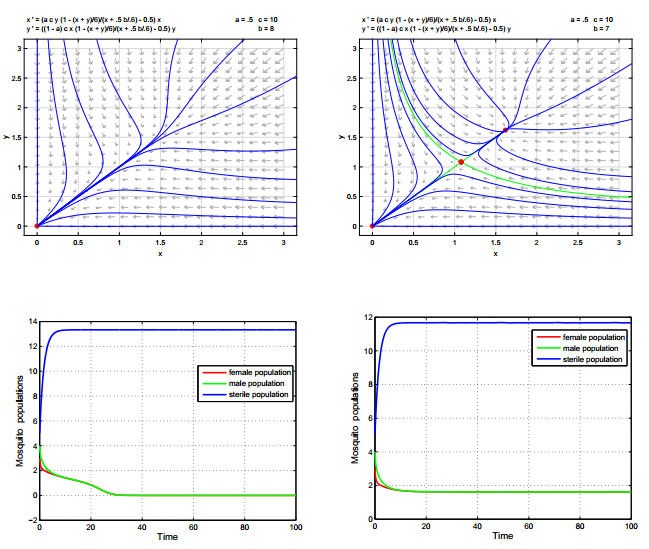
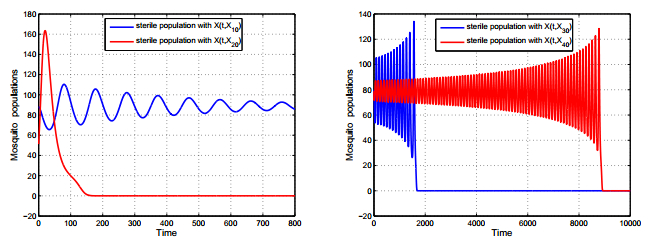
Figures(2)

Shuyang Xue, Meili Li, Junling Ma, Jia Li. Sex-structured wild and sterile mosquito population models with different release strategies[J]. Mathematical Biosciences and Engineering, 2019, 16(3): 1313-1333. doi: 10.3934/mbe.2019064







 DownLoad:
DownLoad: