Mosquito-borne diseases are threatening half of the world's population. To prevent the spread of malaria, dengue fever, or other mosquito-borne diseases, a new disease control strategy is to reduce or eradicate the wild mosquito population by releasing sterile mosquitoes. To study the effects of sterile insect technique on mosquito populations, we developed a mathematical model of constant release of sterile Aedes aegypti mosquitoes with strong and weak Allee effect and considered interspecific competition with Anopheles mosquitoes. We calculated multiple release thresholds and investigated the dynamical behavior of this model. In order to get closer to reality, an impulsive differential equation model was also introduced to study mosquito suppression dynamics under the strategy of releasing $ c $ sterile male mosquitoes at each interval time $ T $. Finally, the relationship between the releasing amount or the waiting period and the number of days required to suppress mosquitoes was illustrated by numerical simulations.
Citation: Chen Liang, Hai-Feng Huo, Hong Xiang. Modelling mosquito population suppression based on competition system with strong and weak Allee effect[J]. Mathematical Biosciences and Engineering, 2024, 21(4): 5227-5249. doi: 10.3934/mbe.2024231
Mosquito-borne diseases are threatening half of the world's population. To prevent the spread of malaria, dengue fever, or other mosquito-borne diseases, a new disease control strategy is to reduce or eradicate the wild mosquito population by releasing sterile mosquitoes. To study the effects of sterile insect technique on mosquito populations, we developed a mathematical model of constant release of sterile Aedes aegypti mosquitoes with strong and weak Allee effect and considered interspecific competition with Anopheles mosquitoes. We calculated multiple release thresholds and investigated the dynamical behavior of this model. In order to get closer to reality, an impulsive differential equation model was also introduced to study mosquito suppression dynamics under the strategy of releasing $ c $ sterile male mosquitoes at each interval time $ T $. Finally, the relationship between the releasing amount or the waiting period and the number of days required to suppress mosquitoes was illustrated by numerical simulations.

| [1] |
S. Ai, J. Li, J. Lu, Mosquito-stage-structured malaria models and their global dynamics, SIAM J. Appl. Math., 72 (2012), 1213–1237. https://doi.org/10.1137/11086031 doi: 10.1137/11086031

|
| [2] |
T. P. Monath, Dengue: the risk to developed and developing countries, Proc. Natl. Acad. Sci. U.S.A., 91 (1994), 2395–2400. https://doi.org/10.1073/pnas.91.7.239 doi: 10.1073/pnas.91.7.239
|
| [3] |
A. Maiti, B. Patra, G. Samanta, Sterile insect release method as a control measure of insect pests: a mathematical model, J. Appl. Math. Comput., 22 (2006), 71–86. https://doi.org/10.1007/BF02832038 doi: 10.1007/BF02832038
|
| [4] |
P. Somwang, J. Yanola, W. Suwan, C. Walton, N. Lumjuan, L. Prapanthadara, et al., Enzymes-based resistant mechanism in pyrethroid resistant and susceptible aedes aegypti strains from northern thailand, Parasitol. Res., 109 (2011), 531–537. https://doi.org/10.1137/11086031 doi: 10.1137/11086031
|
| [5] |
L. Cai, S. Ai, J. Li, Dynamics of mosquitoes populations with different strategies for releasing sterile mosquitoes, SIAM J. Appl. Math., 74 (2014), 1786–1809. https://doi.org/10.1137/13094102X doi: 10.1137/13094102X
|
| [6] |
K. Bourtzis, M. J. Vreysen, Sterile insect technique (sit) and its applications, Insects, 12 (2021), 638. https://doi.org/10.3390/insects12070638 doi: 10.3390/insects12070638
|
| [7] | M. Rafikov, A. Wyse, L. Bevilacqua, Controlling the interaction between wild and transgenic mosquitoes, J. Nonlinear Syst. Appl., 1 (2010), 27–31. |
| [8] |
L. Alphey, M. Benedict, R. Bellini, G. G. Clark, D. A. Dame, M. W. Service, et al., Sterile-insect methods for control of mosquito-borne diseases: an analysis, Vector-Borne Zoonotic Dis., 10 (2010), 295–311. https://doi.org/10.1089/vbz.2009.0014 doi: 10.1089/vbz.2009.0014
|
| [9] |
H. J. Barclay, Pest population stability under sterile releases, Res. Popul. Ecol., 24 (1982), 405–416. https://doi.org/10.1007/BF02515585 doi: 10.1007/BF02515585
|
| [10] |
H. Barclay, M. Mackauer, The sterile insect release method for pest control: a density-dependent model, Environ. Entomol., 9 (1980), 810–817. https://doi.org/10.1093/ee/9.6.810 doi: 10.1093/ee/9.6.810
|
| [11] |
L. Esteva, H. M. Yang, Mathematical model to assess the control of aedes aegypti mosquitoes by the sterile insect technique, Math. Biosci., 198 (2005), 132–147. https://doi.org/10.1016/j.mbs.2005.06.004 doi: 10.1016/j.mbs.2005.06.004
|
| [12] |
J. Flores, A mathematical model for wild and sterile species in competition: Immigration, Physica A, 328 (2003), 214–244. https://doi.org/10.1016/S0378-4371(03)00545-4 doi: 10.1016/S0378-4371(03)00545-4
|
| [13] |
J. Li, Simple mathematical models for interacting wild and transgenic mosquito populations, Math. Biosci., 189 (2004), 39–59. https://doi.org/10.1016/j.mbs.2004.01.001 doi: 10.1016/j.mbs.2004.01.001
|
| [14] |
J. Li, Differential equations models for interacting wild and transgenic mosquito populations, J. Biol. Dyn., 2 (2008), 241–258. https://doi.org/10.1080/17513750701779633 doi: 10.1080/17513750701779633
|
| [15] |
J. Yu, Existence and stability of a unique and exact two periodic orbits for an interactive wild and sterile mosquito model, J. Differ. Equations, 269 (2020), 10395–10415. https://doi.org/10.1016/j.jde.2020.07.019 doi: 10.1016/j.jde.2020.07.019
|
| [16] |
J. Li, L. Cai, Y. Li, Stage-structured wild and sterile mosquito population models and their dynamics, J. Biol. Dyn., 11 (2017), 79–101. https://doi.org/10.1080/17513758.2016.1159740 doi: 10.1080/17513758.2016.1159740
|
| [17] |
J. Li, Modeling of mosquitoes with dominant or recessive transgenes and allee effects, Math. Biosci. Eng., 7 (2010), 101–123. https://doi.org/10.3934/mbe.2010.7.99 doi: 10.3934/mbe.2010.7.99
|
| [18] |
J. Li, Discrete-time models with mosquitoes carrying genetically-modified bacteria, Math. Biosci., 240 (2012), 35–44. https://doi.org/10.1016/j.mbs.2012.05.012 doi: 10.1016/j.mbs.2012.05.012
|
| [19] |
J. Li, New revised simple models for interactive wild and sterile mosquito populations and their dynamics, J. Biol. Dyn., 11 (2017), 316–333. https://doi.org/10.1080/17513758.2016.1216613 doi: 10.1080/17513758.2016.1216613
|
| [20] |
J. Yu, Modeling mosquito population suppression based on delay differential equations, SIAM J. Appl. Math., 78 (2018), 3168–3187. https://doi.org/10.1137/18M1204917 doi: 10.1137/18M1204917
|
| [21] |
B. Zheng, J. Li, J. Yu, Existence and stability of periodic solutions in a mosquito population suppression model with time delay, J. Differ. Equations, 315 (2022), 159–178. https://doi.org/10.1016/j.jde.2022.01.036 doi: 10.1016/j.jde.2022.01.036
|
| [22] |
J. Yu, J. Li, Global asymptotic stability in an interactive wild and sterile mosquito model, J. Differ. Equations, 269 (2020), 6193–6215. https://doi.org/10.1016/j.jde.2020.04.036 doi: 10.1016/j.jde.2020.04.036
|
| [23] |
J. Ito, A. Ghosh, L. A. Moreira, E. A. Wimmer, M. Jacobs-Lorena, Transgenic anopheline mosquitoes impaired in transmission of a malaria parasite, Nature, 417 (2002), 452–455. https://doi.org/10.1038/417452a doi: 10.1038/417452a
|
| [24] |
C. Li, M. T. Marrelli, G. Yan, M. Jacobs-Lorena, Fitness of transgenic anopheles stephensi mosquitoes expressing the sm1 peptide under the control of a vitellogenin promoter, J. Hered., 99 (2008), 275–282. https://doi.org/10.1093/jhered/esn004 doi: 10.1093/jhered/esn004
|
| [25] |
M. Q. Benedict, A. S. Robinson, The first releases of transgenic mosquitoes: an argument for the sterile insect technique, Trends Parasiltol., 19 (2003), 349–355. https://doi.org/10.1016/S1471-4922(03)00144-2 doi: 10.1016/S1471-4922(03)00144-2
|
| [26] |
L. F. Gordillo, Pest persistence and eradication conditions in a deterministic model for sterile insect release, J. Biol. Dyn., 9 (2015), 64–78. https://doi.org/10.1080/17513758.2014.942393 doi: 10.1080/17513758.2014.942393
|
| [27] |
Y. Dumont, J. Tchuenche, Mathematical studies on the sterile insect technique for the chikungunya disease and aedes albopictus, J. Math. Biol., 65 (2012), 809–854. https://doi.org/10.1007/s00285-011-0477-6 doi: 10.1007/s00285-011-0477-6
|
| [28] |
M. Huang, X. Song, J. Li, Modelling and analysis of impulsive releases of sterile mosquitoes, J. Biol. Dyn., 11 (2017), 147–171. https://doi.org/10.1080/17513758.2016.1254286 doi: 10.1080/17513758.2016.1254286
|
| [29] |
L. Cai, S. Ai, G. Fan, Dynamics of delayed mosquitoes populations models with two different strategies of releasing sterile mosquitoes, Math. Biosci. Eng., 15 (2018), 1181–1202. https://doi.org/10.3934/mbe.2018054 doi: 10.3934/mbe.2018054
|
| [30] | W. C. Allee, The Social Life of Animals, W. W. Norton and Company, New York, 1938. https://doi.org/10.5962/bhl.title.7226 |
| [31] |
M. J. Groom, Allee effects limit population viability of an annual plant, Am. Nat., 151 (1998), 487–496. https://doi.org/10.1086/286135 doi: 10.1086/286135
|
| [32] | R. W. Craig, J. L. Katherine, J. W. Natasha, R. S. Veronica, The allee effect in site choice behaviour of egg-laying dengue vector mosquitoes, Trop. Biomed., 25 (2008), 140–144. |
| [33] |
M. Strugarek, H. Bossin, Y. Dumont, On the use of the sterile insect release technique to reduce or eliminate mosquito populations, Appl. Math. Modell., 68 (2019), 443–470. https://doi.org/10.1016/j.apm.2018.11.026 doi: 10.1016/j.apm.2018.11.026
|
| [34] |
X. Song, Z. Xiang, The prey-dependent consumption two-prey one-predator models with stage structure for the predator and impulsive effects, J. Theor. Biol., 242 (2006), 683–698. https://doi.org/10.1016/j.jtbi.2006.05.002 doi: 10.1016/j.jtbi.2006.05.002
|
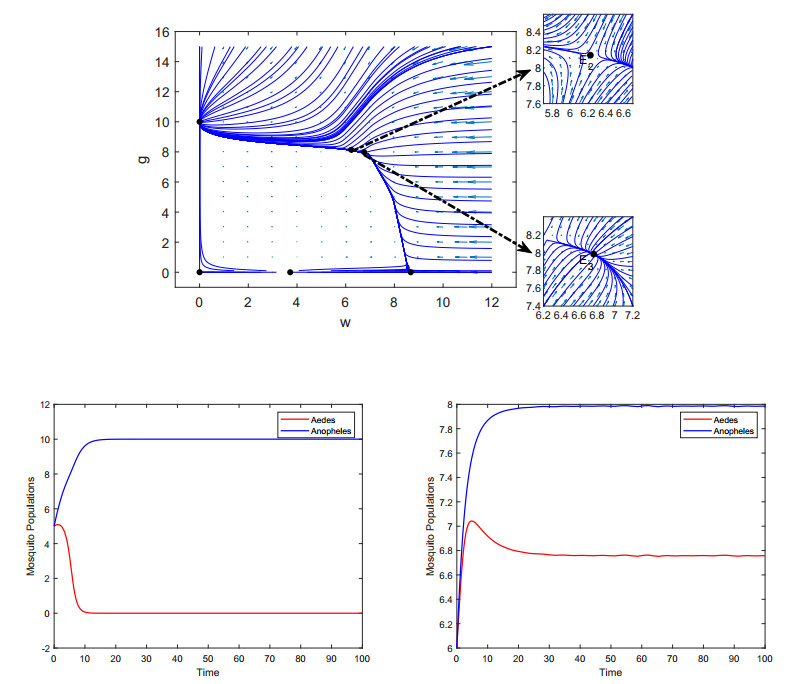








Figures(9)

Chen Liang, Hai-Feng Huo, Hong Xiang. Modelling mosquito population suppression based on competition system with strong and weak Allee effect[J]. Mathematical Biosciences and Engineering, 2024, 21(4): 5227-5249. doi: 10.3934/mbe.2024231







 DownLoad:
DownLoad: