Hepatitis B is one of the global health issues caused by the hepatitis B virus (HBV), producing 1.1 million deaths yearly. The acute and chronic phases of HBV are significant because worldwide, approximately 250 million people are infected by chronic hepatitis B. The chronic stage is a long-term, persistent infection that can cause liver damage and increase the risk of liver cancer. In the case of multiple phases of infection, a generalized saturated incidence rate model is more reasonable than a simply saturated incidence because it captures the complex dynamics of the different infection phases. In contrast, a simple saturated incidence rate model assumes a fixed shape for the incidence rate curve, which may not accurately reflect the dynamics of multiple infection phases. Considering HBV and its various phases, we constructed a model to present the dynamics and control strategies using the generalized saturated incidence. First, we proved that the model is well-posed. We then found the reproduction quantity and model equilibria to discuss the time dynamics of the model and investigate the conditions for stabilities. We also examined a control mechanism by introducing various controls to the model with the aim to increase the population of those recovered and minimize the infected people. We performed numerical experiments to check the biological significance and control implementation.
Citation: Tahir Khan, Fathalla A. Rihan, Muhammad Ibrahim, Shuo Li, Atif M. Alamri, Salman A. AlQahtani. Modeling different infectious phases of hepatitis B with generalized saturated incidence: An analysis and control[J]. Mathematical Biosciences and Engineering, 2024, 21(4): 5207-5226. doi: 10.3934/mbe.2024230
Hepatitis B is one of the global health issues caused by the hepatitis B virus (HBV), producing 1.1 million deaths yearly. The acute and chronic phases of HBV are significant because worldwide, approximately 250 million people are infected by chronic hepatitis B. The chronic stage is a long-term, persistent infection that can cause liver damage and increase the risk of liver cancer. In the case of multiple phases of infection, a generalized saturated incidence rate model is more reasonable than a simply saturated incidence because it captures the complex dynamics of the different infection phases. In contrast, a simple saturated incidence rate model assumes a fixed shape for the incidence rate curve, which may not accurately reflect the dynamics of multiple infection phases. Considering HBV and its various phases, we constructed a model to present the dynamics and control strategies using the generalized saturated incidence. First, we proved that the model is well-posed. We then found the reproduction quantity and model equilibria to discuss the time dynamics of the model and investigate the conditions for stabilities. We also examined a control mechanism by introducing various controls to the model with the aim to increase the population of those recovered and minimize the infected people. We performed numerical experiments to check the biological significance and control implementation.

| [1] |
M. H. Chang, Hepatitis B virus infection, Semin. Fetal Neonatal Med., 12 (2007), 160–167. https://doi.org/10.1017/CBO9781139012102 doi: 10.1017/CBO9781139012102

|
| [2] |
M. R. Hall, D. Ray, J. A. Payne, Prevalence of hepatitis C, hepatitis B, and human immunodeficiency virus in a grand rapids, michigan emergency department, J. Emerg. Med., 38 (2010), 401–405. https://doi.org/10.1016/j.jemermed.2008.03.036 doi: 10.1016/j.jemermed.2008.03.036
|
| [3] |
W. Edmunds, G. Medley, D. Nokes, A. Hall, H. Whittle, The influence of age on the development of the hepatitis B carrier state, Proc. R. Soc. Ser. B Biol. Sci., 253 (1993), 197–201. https://doi.org/10.1098/rspb.1993.0102 doi: 10.1098/rspb.1993.0102
|
| [4] |
J. Mann, M. Roberts, Modelling the epidemiology of hepatitis B in New Zealand, J. Theor. Biol., 269 (2011), 266–272. https://doi.org/10.1016/j.jtbi.2010.10.028 doi: 10.1016/j.jtbi.2010.10.028
|
| [5] | M. Jakab, J. Farrington, L. Borgermans, F. Mantingh, Health Systems Respond to Noncommunicable Diseases: Time for Ambition, World Health Organization, Regional Office for Europe, 2018. |
| [6] | D. Lavanchy, Hepatitis B virus epidemiology, disease burden, treatment, and current and emerging prevention and control measures, J. Viral Hepatitis, 11 (2004), 97–107. |
| [7] |
B. J. McMahon, Epidemiology and natural history of hepatitis B, Semin. Liver Dis., 25 (2005), 3–8. https://doi.org/10.1055/s-2005-915644 doi: 10.1055/s-2005-915644
|
| [8] |
F. Brauer, Some simple epidemic models, Math. Biosci. Eng., 3 (2006). https://doi.org/10.3934/mbe.2006.3.1 doi: 10.3934/mbe.2006.3.1
|
| [9] |
J. Wang, J. Pang, X. Liu, Modelling diseases with relapse and nonlinear incidence of infection: a multi-group epidemic model, J. Biol. Dyn., 8 (2014), 99–116. https://doi.org/10.1080/17513758.2014.912682 doi: 10.1080/17513758.2014.912682
|
| [10] |
J. Wang, R. Zhang, T. Kuniya, The stability analysis of an SVEIR model with continuous age-structure in the exposed and infectious classes, J. Biol. Dyn., 9 (2015), 73–101. https://doi.org/10.1080/17513758.2015.1006696 doi: 10.1080/17513758.2015.1006696
|
| [11] |
B. Alten, C. Maia, M. O. Afonso, L. Campino, M. Jiménez, E. González, et al., Seasonal dynamics of phlebotomine sand fly species proven vectors of mediterranean leishmaniasis caused by leishmania infantum, PLoS Negl. Trop. Dis., 10 (2016), e0004458. https://doi.org/10.1371/journal.pntd.0004458 doi: 10.1371/journal.pntd.0004458
|
| [12] |
D. Sereno, Epidemiology of vector-borne diseases 2.0, Microorganisms, 10 (2022), 1555. https://doi.org/10.3390/microorganisms10081555 doi: 10.3390/microorganisms10081555
|
| [13] |
B. Li, H. Liang, L. Shi, Q. He, Complex dynamics of Kopel model with nonsymmetric response between oligopolists, Chaos, Solitons Fractals, 156 (2022), 111860. https://doi.org/10.1016/j.chaos.2022.111860 doi: 10.1016/j.chaos.2022.111860
|
| [14] |
Q. He, M. U. Rahman, C. Xie, Information overflow between monetary policy transparency and inflation expectations using multivariate stochastic volatility models, Appl. Math. Sci. Eng., 31 (2023), 2253968. https://doi.org/10.1080/27690911.2023.2253968 doi: 10.1080/27690911.2023.2253968
|
| [15] |
B. Li, T. Zhang, C. Zhang, Investigation of financial bubble mathematical model under fractal-fractional Caputo derivative, Fractals, 31 (2023), 1–13. https://doi.org/10.1142/S0218348X23500500 doi: 10.1142/S0218348X23500500
|
| [16] | F. Brauer, C. Castillo-Chavez, C. Castillo-Chavez, Mathematical Models in Population Biology and Epidemiology, Springer, 2 (2012). |
| [17] |
W. O. Kermack, A. G. McKendrick, A contribution to the mathematical theory of epidemics, Proc. R. Soc. London, Ser. A, 115 (1927), 700–721. https://doi.org/10.1098/rspa.1927.0118 doi: 10.1098/rspa.1927.0118
|
| [18] |
X. Zhai, W. Li, F. Wei, X. Mao, Dynamics of an HIV/AIDS transmission model with protection awareness and fluctuations, Chaos, Solitons Fractals, 169 (2023), 113224. https://doi.org/10.1016/j.chaos.2023.113224 doi: 10.1016/j.chaos.2023.113224
|
| [19] |
J. Williams, D. Nokes, G. Medley, R. Anderson, The transmission dynamics of hepatitis B in the UK: a mathematical model for evaluating costs and effectiveness of immunization programmes, Epidemiol. Infect., 116 (1996), 71–89. https://doi.org/10.1017/S0950268800058970 doi: 10.1017/S0950268800058970
|
| [20] |
F. A. Rihan, H. J. Alsakaji, Analysis of a stochastic hbv infection model with delayed immune response, Math. Biosci. Eng., 18 (2021), 5194–5220. https://doi.org/10.3934/mbe.2021264 doi: 10.3934/mbe.2021264
|
| [21] |
T. Xue, L. Zhang, X. Fan, Dynamic modeling and analysis of hepatitis B epidemic with general incidence, Math. Biosci. Eng., 20 (2023), 10883–10908. https://doi.org/10.3934/mbe.2023483 doi: 10.3934/mbe.2023483
|
| [22] |
G. F. Medley, N. A. Lindop, W. J. Edmunds, D. J. Nokes, Hepatitis-B virus endemicity: heterogeneity, catastrophic dynamics and control, Nat. Med., 7 (2001), 619–624. https://doi.org/10.1038/87953 doi: 10.1038/87953
|
| [23] |
S. Zhao, Z. Xu, Y. Lu, A mathematical model of hepatitis B virus transmission and its application for vaccination strategy in China, Int. J. Epidemiol., 29 (2000), 744–752. https://doi.org/10.1093/ije/29.4.744 doi: 10.1093/ije/29.4.744
|
| [24] |
T. Khan, G. Zaman, M. I. Chohan, The transmission dynamic of different hepatitis B-infected individuals with the effect of hospitalization, J. Biol. Dyn., 12 (2018), 611–631. https://doi.org/10.1080/17513758.2018.1500649 doi: 10.1080/17513758.2018.1500649
|
| [25] |
T. Khan, Z. Ullah, N. Ali, G. Zaman, Modeling and control of the hepatitis B virus spreading using an epidemic model, Chaos, Solitons Fractals, 124 (2019), 1–9. https://doi.org/10.1016/j.chaos.2019.04.033 doi: 10.1016/j.chaos.2019.04.033
|
| [26] |
M. Fan, M. Y. Li, K. Wang, Global stability of an SEIS epidemic model with recruitment and a varying total population size, Math. Biosci., 170 (2001), 199–208. https://doi.org/10.1016/S0025-5564(00)00067-5 doi: 10.1016/S0025-5564(00)00067-5
|
| [27] |
J. Li, Z. Ma, Qualitative analyses of SIS epidemic model with vaccination and varying total population size, Math. Comput. Modell., 35 (2002), 1235–1243. https://doi.org/10.1016/S0895-7177(02)00082-1 doi: 10.1016/S0895-7177(02)00082-1
|
| [28] |
L. Zou, W. Zhang, S. Ruan, Modeling the transmission dynamics and control of hepatitis B virus in China, J. Theor. Biol., 262 (2010), 330–338. https://doi.org/10.1016/j.jtbi.2009.09.035 doi: 10.1016/j.jtbi.2009.09.035
|
| [29] |
V. Capasso, G. Serio, A generalization of the kermack-mckendrick deterministic epidemic model, Math. Biosci., 42 (1978), 43–61. https://doi.org/10.1016/0025-5564(78)90006-8 doi: 10.1016/0025-5564(78)90006-8
|
| [30] |
J. Zhang, J. Jia, X. Song, Analysis of an SEIR epidemic model with saturated incidence and saturated treatment function, Sci. World J., 2014 (2014). https://doi.org/10.1155/2014/910421 doi: 10.1155/2014/910421
|
| [31] |
T. Khan, G. Zaman, Classification of different hepatitis B infected individuals with saturated incidence rate, SpringerPlus, 5 (2016), 1–16. https://doi.org/10.1186/s40064-016-2706-3 doi: 10.1186/s40064-016-2706-3
|
| [32] |
D. Li, F. Wei, X. Mao, Stationary distribution and density function of a stochastic SVIR epidemic model, J. Franklin Inst., 359 (2022), 9422–9449. https://doi.org/10.1016/j.jfranklin.2022.09.026 doi: 10.1016/j.jfranklin.2022.09.026
|
| [33] |
O. Diekmann, J. A. P. Heesterbeek, J. A. Metz, On the definition and the computation of the basic reproduction ratio $r_0$ in models for infectious diseases in heterogeneous populations, J. Math. Biol., 28 (1990), 365–382. https://doi.org/10.1007/BF00178324 doi: 10.1007/BF00178324
|
| [34] |
P. Van den Driessche, J. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180 (2002), 29–48. https://doi.org/10.1016/S0025-5564(02)00108-6 doi: 10.1016/S0025-5564(02)00108-6
|
| [35] |
A. V. Kamyad, R. Akbari, A. A. Heydari, A. Heydari, Mathematical modeling of transmission dynamics and optimal control of vaccination and treatment for hepatitis B virus, Comput. Math. Methods Med., 2014 (2014). https://doi.org/10.1155/2014/475451 doi: 10.1155/2014/475451
|
| [36] |
G. Zaman, Y. H. Kang, I. H. Jung, Stability analysis and optimal vaccination of an SIR epidemic model, BioSystems, 93 (2008), 240–249. https://doi.org/10.1016/j.biosystems.2008.05.004 doi: 10.1016/j.biosystems.2008.05.004
|
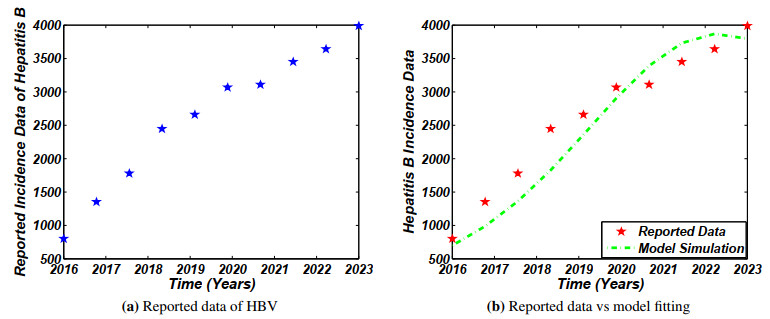



Figures(4)

Tahir Khan, Fathalla A. Rihan, Muhammad Ibrahim, Shuo Li, Atif M. Alamri, Salman A. AlQahtani. Modeling different infectious phases of hepatitis B with generalized saturated incidence: An analysis and control[J]. Mathematical Biosciences and Engineering, 2024, 21(4): 5207-5226. doi: 10.3934/mbe.2024230







 DownLoad:
DownLoad: