Citation: Yuanxian Hui, Genghong Lin, Qiwen Sun. Oscillation threshold for a mosquito population suppression model with time delay[J]. Mathematical Biosciences and Engineering, 2019, 16(6): 7362-7374. doi: 10.3934/mbe.2019367

| [1] | L. Alphey, M. Benedict, R. Bellini, et al., Sterile-insect methods for control of mosquito-borne disease: An analysis, Vector Borne Zoonotic Dis., 10 (2010), 295–311. |
| [2] | R. S. Patterson, D. E. Weidhaass, H. R. Ford, et al., Suppression and elimination of an island population of Culex pipiens quinquefasciatus with sterile males, Science, 168 (1970), 1368–1369. |
| [3] | S. Ai, J. Li and J. Lu, Mosquito-stage-structured malaria models and their global dynamics, SIAM J. Appl. Math., 72 (2012), 1213–1237. |
| [4] | L. Cai, S. Ai and J. Li, Dynamics of mosquitoes populations with differential strategies for releas-ing sterile mosquitoes, SIAM J. Appl. Math., 74 (2014), 1786–1809. |
| [5] | K. R. Fister, M. L. Mccarthy, S. F. Oppenheimer, et al., Optimal control of insects through sterile insect release and habitat modification, Math. Biosci., 244 (2013), 201–212. |
| [6] | J. Li, L. Cai and Y. Li, Stage-structured wild and sterile mosquito population models and their dynamics, J. Biol. Dynam., 11 (2017), 79–101. |
| [7] | J. Li, New revised simple models for interactive wild and sterile mosquito populations and their dynamics, J. Biol. Dynam., 11 (2017), 316–333. |
| [8] | X. Zhang, S. Tang and Q. Liu, Models to assess the effects of non-identical sex ratio augmentations of Wolbachia-carrying mosquitoes on the control of dengue disease, Math. Biosci., 299 (2018), 58–72. |
| [9] | B. Zheng, M. Tang, J. Yu, et al., Wolbachia spreading dynamics in mosquitoes with imperfect maternal transmission, J. Math. Biol., 76 (2018), 235–263. |
| [10] | L. Cai, S. Ai and G. Fan, Dynamics of delayed mosquitoes populations with two different strategies of releasing sterile mosquitoes, Math. Biosci. Eng., 15 (2018), 1181–1202. |
| [11] | M. Huang, J. Luo, L. Hu, et al., Assessing the efficiency of Wolbachia driven Aedes mosquito suppression by delay differential equations, J. Theor. Biol., 440 (2018), 1–11. |
| [12] | J. Yu, Modelling mosquito population suppression based on delay differential equations, SIAM J. Appl. Math., 78 (2018), 3168–3187. |
| [13] | B. Zheng, M. Tang and J. Yu, Modeling Wolbachia spread in mosquitoes through delay differential equations, SIAM J. Appl. Math., 74 (2014), 743–770. |
| [14] | M. Huang, M. Tang, J. Yu, et al., The impact of mating competitiveness and incomplete cytoplas-mic incompatibility on Wolbachia-driven mosquito population suppression, Math. Biosci. Eng., 16 (2019), 4741–4757. |
| [15] | H. Diaz, A. A. Ramirez, A. Olarte, et al., A model for the control of malaria using genetically modified vectors, J. Theor. Biol., 276 (2011), 57–66. |
| [16] | K. R. Fister, M. L. Mccarthy and S. F. Oppenheimer, Diffusing wild type and sterile mosquitoes in an optimal control setting. Optimal control of insects through sterile insect release and habitat modification, Math. Biosci., 302 (2018), 100–115. |
| [17] | M. Huang, M. Tang and J. Yu, Wolbachiainfection dynamics by reaction-diffusion equations, Sci. China Math., 58 (2015), 77–96. |
| [18] | M. Huang, J. Yu, L. Hu, et al., Qualitative analysis for a Wolbachia infection model with diffusion, Sci. China Math., 59 (2016), 1249–1266. |
| [19] | J. C. Floresa, A mathematical model for wild and sterile species in competition: Immigration, Phys. A., 328 (2003), 214–224. |
| [20] | L. Hu, M. Tang, Z. Wu, et al., The threshold infection level for Wolbachia invasion in random environment, J. Differ. Equations, 266 (2019), 4377–4393. |
| [21] | H. L. Smith, An introduction to delay differential equations with applications to the life sciences, Springer, 2011. |
| [22] | I. Györi and G. Ladas, Oscillation theory of delay differential equations: with application, Oxford University Press, 1991. |
| [23] | R. Anguelova, Y. Dumontb and J. Lubuma, Mathematical modeling of sterile insect technology for control of Anopheles mosquito, Comput. Math. Appl., 64 (2012), 374–389. |
| [24] | N. Chitnis, J. M. Hyman and J. M. Cushing, Determining important parameters in the spread through the sensitivity analysis of a mathematical model, Bull. Math. Biol., 70 (2008), 1272–1296. |
| [25] | A. A. Hoffmann, M. Turelli and L. G. Harshman, Factors affecting the distribution of cytoplasmic incompatibility in Drosophia simulans, Genetics, 126 (1990), 933–948. |
| [26] | Y. Li, F. Kamara, G. Zhou, et al., Urbanization increases Aedes albopictus larval habitats and accelerates mosquito development and survivorship, PLoS Negl. Trop. Dis., 8 (2014), e3301. |
| [27] | F. Liu, C. Yao, P. Lin, et al., Studies on life table of the nature population of Aedes albopictus, Acta Sci. Natur. Univ. Sunyatseni, 31 (1992), 84–93. |
| [28] | Z. Liu, Y. Zhang and Y. Yang, Population dynamics of Aedes (stegomyia) albopictus (Skuse) under laboratory conditions, Acta Entomol. Sin., 28 (1985), 274–280. |
| [29] | D. Zhang, X. Zheng, Z. Xi, et al., Combining the sterile insect technique with the incompatible insect technique: I-impact of Wolbachia infection on the fitness of triple- and double-infected strains of Aedes albopictus, PLoS One, 10 (2015), 1–13. |
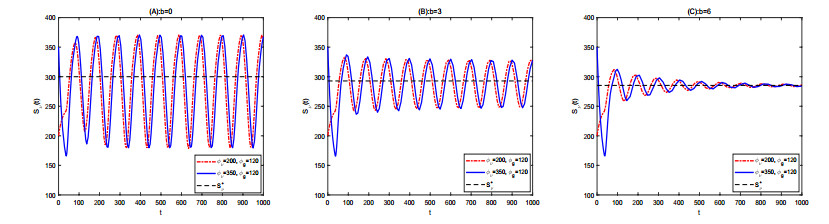
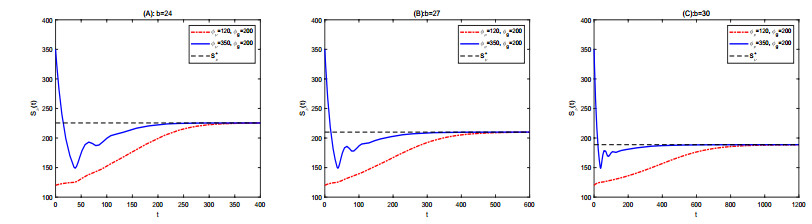
Figures(2) / Tables(1)

Yuanxian Hui, Genghong Lin, Qiwen Sun. Oscillation threshold for a mosquito population suppression model with time delay[J]. Mathematical Biosciences and Engineering, 2019, 16(6): 7362-7374. doi: 10.3934/mbe.2019367







 DownLoad:
DownLoad: