Citation: Xiaoli Wang, Junping Shi, Guohong Zhang. Bifurcation analysis of a wild and sterile mosquito model[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 3215-3234. doi: 10.3934/mbe.2019160

| [1] | H. Gaff, D. Hartley and N. Leahy, An epidemiological model of rift valley fever, Electron. J. Differ. Eq., 115 (2007), 1–12. |
| [2] | E. Michael, M. Malecela-Lazaro, B. Maegga, et al., Mathematical models and lymphatic filariasis control: monitoring and evaluating interventions, Trends Parasitol., 22 (2006), 529–535. |
| [3] | E. Newton and P. Reiter, A model of the transmission of dengue fever with an evaluation of the impact of ultra-low volume (ULV) insecticide applications on dengue epidemics, Am. J. Trop. Hygiene Med., 47 (1992), 709. |
| [4] | M. Predescu, G. Sirbu, R. Levins, et al., On the dynamics of a deterministic and stochastic model for mosquito control, Appl. Math. Lett., 20 (2007), 919–925. |
| [5] | D. Smith, J. Dushoff and F. McKenzie, The risk of a mosquito-borne infection in a heterogeneous environment, PLOS Biol., 2 (2007), e368. 6. A. Wyse, L. Bevilacqua and M. Rafikov, Simulating malaria model for different treatment intensities in a variable environmen, Ecol. Modell., 206 (2010), 322–330. |
| [6] | 7. L. Cai, S. Ai and J. Li, Dynamics of mosquitoes populations with different strategies for releasing sterile mosquitoes, SIAM J. Appl. Math., 74 (2014), 1781809. |
| [7] | 8. CDC, Epidemiology, 2014. Available from: http://www.cdc.gov/dengue/epidemiology. |
| [8] | 9. S. Buhr, Googles life sciences unit is releasing 20 million bacteria-infected mosquitoes in Fresno, 2017. Available from: https://techcrunch.com/2017/07/14/googles-life-sciences-unit-is-releasing- 20-million-bacteria-infected-mosquitoes-in-fresno/. |
| [9] | 10. H. Barclay, Pest population stability under sterile releases, Res. Popul. Ecol., 24 (12), 405–416. |
| [10] | 11. H. Barclay, Mathematical models for the use of sterile insects, in sterile insect technique, Springer Netherlands, (2005), 147–174. |
| [11] | 12. H. Barclay and M. Mackauer, The sterile insect release method for pest control: a densitydependent model, Environ. Entomol., 9 (1980), 810–817. |
| [12] | 13. L. Fister, M. McCarthy, S. Oppenheimer, et al., Optimal control of insects through sterile insect release and habitat modification, Math. Biosci., 244 (2013), 201–2 |
| [13] | 14. J. Floresa, A mathematical model for wild and sterile species in competition: Immigration, Phys. A, 328 (2003), 214–224. |
| [14] | 15. J. Li, New revised simple models for interactive wild and sterile mosquito populations and their dynamics, J. Biol. Dyn., 11 (2017), 316–313. |
| [15] | 16. J. Li, M. Han and J. Yu, Simple paratransgenic mosquitoes models and their dynamics, Math. Biosci., 306 (2018), 20–31. |
| [16] | 17. J. Li and Z. Yuan, Modelling releases of sterile mosquitoes with different strategies, J. Biol. Dyn., 9 (2015), 1–14. |
| [17] | 18. B. Zheng, W. Guo, L. Hu, et al., Complex Wolbachia infection dynamics in mosquitoes with imperfect maternal transmission, Math. Biosci. Eng., 15 (2018), 523–541. |
| [18] | 19. B. Zheng, M. X. Tang, J. Yu, et al., Wolbachia spreading dynamics in mosquitoes imperfect maternal transmission, J. Math. Biol., 76 (20, 235–263. |
| [19] | 20. B. Zheng and J. Yu, Characterization of Wolbachia enhancing domain in mosquitoes with imperfect maternal transmission, J. Biol. Dyn., 12 (2018), 596–610. |
| [20] | 21. S. Ai, J. Li and J. Lu, Mosquito-stage-structured malaria models and their global dynamics, SIAM J. Appl. Math., 72 (4), 1213–1237. |
| [21] | 22. J. Li and L. Cai, Stage-structured wild and sterile mosquito population models and their dynamics, J. Biol. Dyn., 11 (2017), 79–101. |
| [22] | 23. M. Huang, J. Luo, L. Hu, et al., Assessing the efficiency of Wolbachia driven Aedes mosquito suppression by delay differential equations, J. Theoret. Biol., 440 (2018), 1–11. |
| [23] | 24. J. Yu, Modeling mosquito population suppression based on delay differential equations, SIAM J. Appl. Math., 78 (2018), 3168–3187. |
| [24] | 25. B. Zheng, M. Tang and J. Yu, ModelingWolbachia spread in mosquitoes through delay differential equations, SIAM J. Appl. Math., 74 (2014), 743–770. |
| [25] | 26. M. Huang, M. Tang and J. Yu, Wolbachia infection dynamics by reaction-diffusion equations, Sci. China. Math., 58 (2015), 77–96. |
| [26] | 27. M. Huang, J. Yu, L. Hu, et al., Qualitative analysis for aWolbachia infection model with diffusion, Sci. China. Math., 59 (2016), 1249–1. |
| [27] | 28. L. Hu, M. Huang, M. Tang, et al., Wolbachia spread dynamics in stochastic environments, Theor. Popul. Biol., 106 (2015), 32–44. |
| [28] | 29. W. Allee, The Social Life of Animals, Heinemann, London, 1938. |
| [29] | 30. S. Schreiber, Allee effects, extinctions, and chaotic transients in simple population models, Theor. Popul. Biol., 64 (2003), 201–209. |
| [30] | 31. T. Sturm, A. Weber, E. Abdel-Rahman, et al., Investigating algebraic and logical algorithma to solve Hopf bifurcation problems in algebraic biology, Math. Comput. Sci., 2 (2009), 493–515. |
| [31] | 32. J. Alexander and J. Yorke, Global bifurcations of periodic orbits, Amer. J. Math., 100 (1978), 263–292. |
| [32] | 33. S. Chow and J. Mallet-Paret, The Fuller index and global Hopf bifurcation, J. Differ. Equations, 29 (1978), 66–85. |
| [33] | 34. X. Wang, J. Shi and G. Zhang, Interaction between water and plants: rich dynamics in a simple model, Discrete Contin. Dyn. Syst. Ser. B, 22 (2017), 2971–3006. |
| [34] | 35. R. Liu, Z. Feng, H. Zhu, et al., Bifurcation analysis of a plant-herbivore model with toxindetermined functional response, J. Differ. Equations, 245 (2008), 442–467. |
| [35] | 36. W. Wang and S. Ruan, Bifurcation in an epidemic model with constant removal rate of the infectives, J. Math. Anal. Appl., 291 (2004), 775–793. |
| [36] | 37. L. Perko, Differential Equations and Dynamical Systems Springer, New York, 1996. |
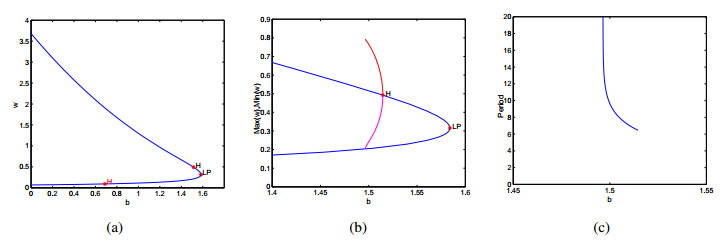
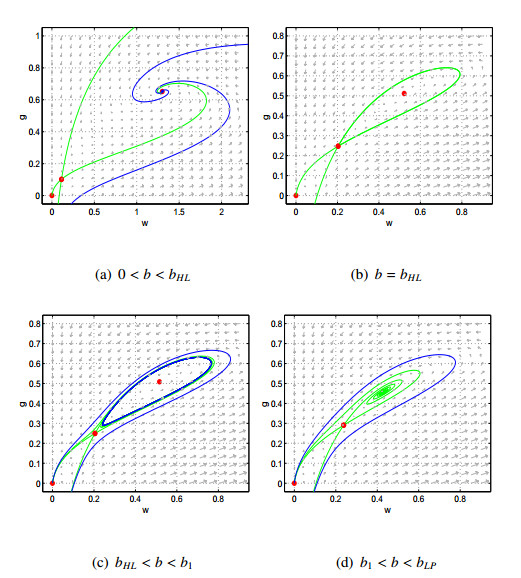
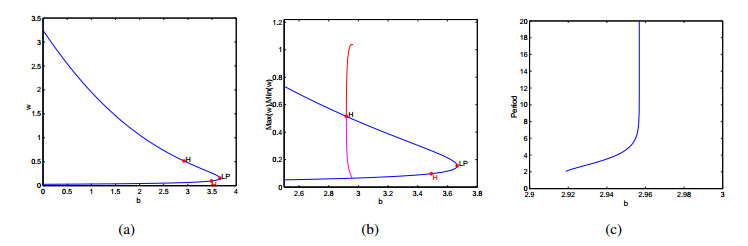
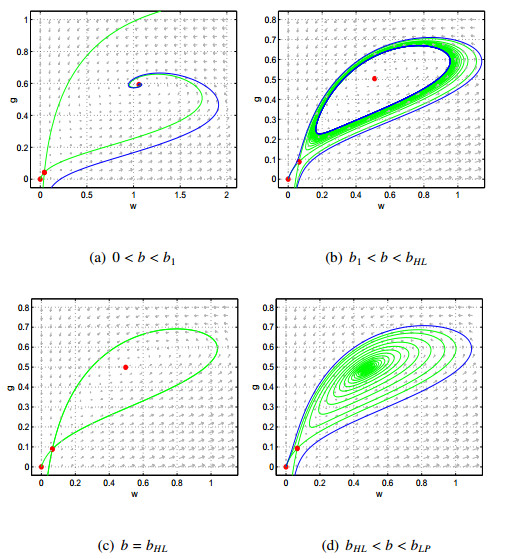
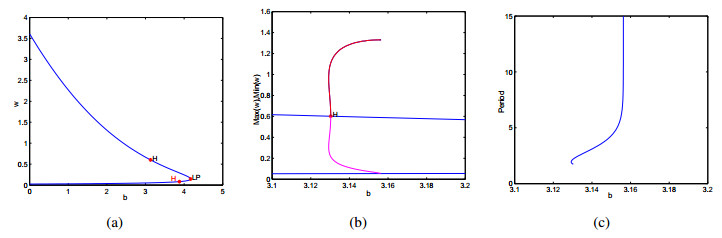

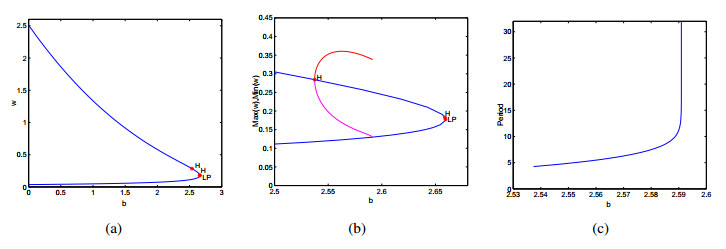
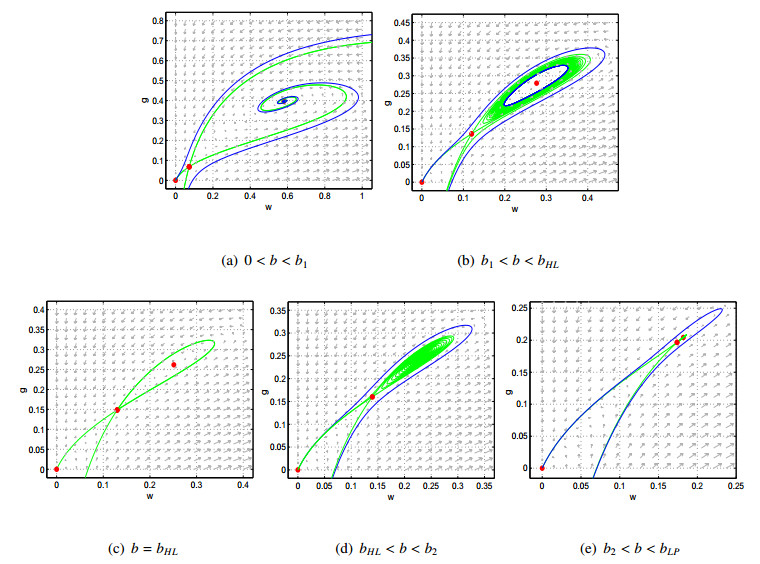
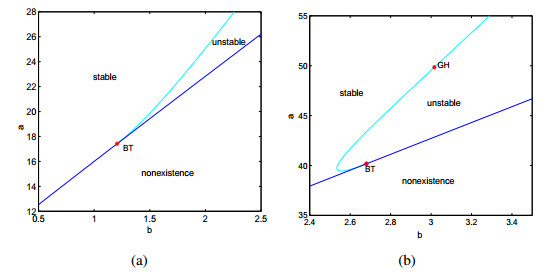
Figures(9)

Xiaoli Wang, Junping Shi, Guohong Zhang. Bifurcation analysis of a wild and sterile mosquito model[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 3215-3234. doi: 10.3934/mbe.2019160







 DownLoad:
DownLoad: