Non-chromosomal structure maintenance protein condensin complex I subunit H (NCAPH) has been reported to play a regulatory role in a variety of cancers and is associated with tumor poor prognosis. This study aims to explore the potential role of NCAPH with a view to providing insights on pathologic mechanisms.
The expression of NCAPH in different tumors was explored by The Cancer Genome Atlas (TCGA) and Genotype Tissue Expression (GTEx). The prognostic value of NCAPH was retrieved through GEPIA and Kaplan-Meier Plotter databases. Tumor Immunity Estimation Resource (TIMER) and Single-Sample Gene Set Enrichment Analysis (GSEA) to search for the association of NCAPH with tumor immune infiltration. The cBioPortal and PhosphoSite Plus databases showed NCAPH phosphorylation status in tumors. Gene set enrichment analysis (GSEA) was performed using bioinformatics.
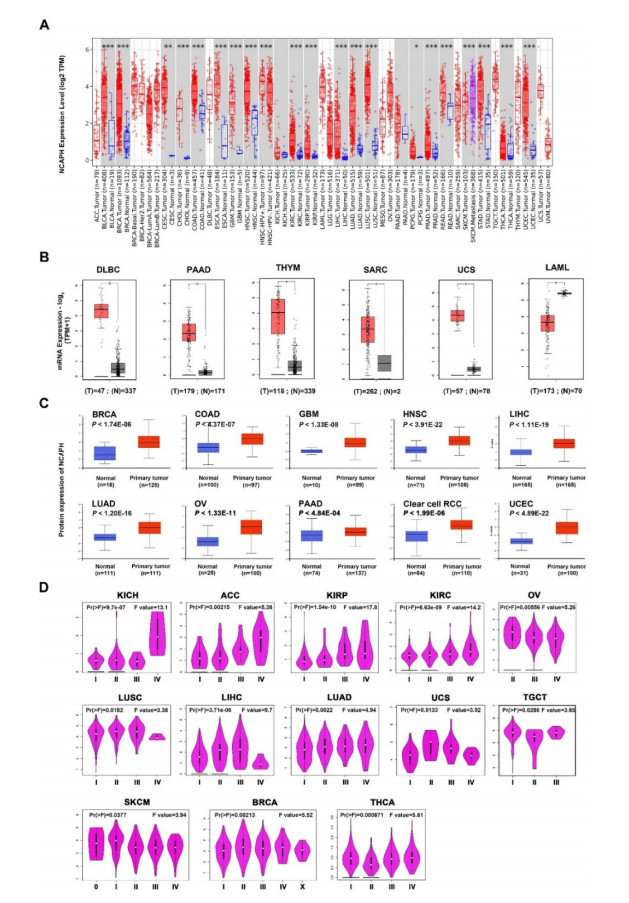
Our findings revealed that NCAPH showed high expression levels in a wide range of tumor types, and was strongly correlated with the prognosis of patients. Moreover, a higher phosphorylation level at S59, S67, S76, S190, S222 and T38 site was discovered in head and neck squamous cell carcinoma (HNSC). NCAPH overexpression was positively correlated with the infiltration level of CD8+T cells and myeloid dendritic infiltration in breast cancer and thymoma.
The up-regulation of NCAPH was significantly correlated with the poor prognosis and immune infiltration in pan-cancer, and NCAPH could be served as a potential immunotherapeutic target for cancers.
Citation: Ying Liu, Xiao Ma, Linyuan Feng, Zhenhua Lin, Xianchun Zhou. An integrative pan-cancer analysis reveals the carcinogenic effects of NCAPH in human cancer[J]. Mathematical Biosciences and Engineering, 2023, 20(1): 76-92. doi: 10.3934/mbe.2023005
Non-chromosomal structure maintenance protein condensin complex I subunit H (NCAPH) has been reported to play a regulatory role in a variety of cancers and is associated with tumor poor prognosis. This study aims to explore the potential role of NCAPH with a view to providing insights on pathologic mechanisms.
The expression of NCAPH in different tumors was explored by The Cancer Genome Atlas (TCGA) and Genotype Tissue Expression (GTEx). The prognostic value of NCAPH was retrieved through GEPIA and Kaplan-Meier Plotter databases. Tumor Immunity Estimation Resource (TIMER) and Single-Sample Gene Set Enrichment Analysis (GSEA) to search for the association of NCAPH with tumor immune infiltration. The cBioPortal and PhosphoSite Plus databases showed NCAPH phosphorylation status in tumors. Gene set enrichment analysis (GSEA) was performed using bioinformatics.
Our findings revealed that NCAPH showed high expression levels in a wide range of tumor types, and was strongly correlated with the prognosis of patients. Moreover, a higher phosphorylation level at S59, S67, S76, S190, S222 and T38 site was discovered in head and neck squamous cell carcinoma (HNSC). NCAPH overexpression was positively correlated with the infiltration level of CD8+T cells and myeloid dendritic infiltration in breast cancer and thymoma.
The up-regulation of NCAPH was significantly correlated with the poor prognosis and immune infiltration in pan-cancer, and NCAPH could be served as a potential immunotherapeutic target for cancers.

| [1] |
T. G. Phan, P. I. Croucher, The dormant cancer cell life cycle, Nat. Rev. Cancer, 20 (2020), 398–411. https://doi.org/10.1038/s41568-020-0263-0 doi: 10.1038/s41568-020-0263-0

|
| [2] |
Y. Nie, X. Wang, F. Yang, Z. Zhou, J. Wang, K. Chen, Surgical prognosis of synchronous multiple primary lung cancer: Systematic review and meta-analysis, Clin. Lung Cancer, 22 (2021), 341–350.e3. https://doi.org/10.1016/j.cllc.2020.10.022 doi: 10.1016/j.cllc.2020.10.022
|
| [3] |
T. J. Willenbrink, E. S. Ruiz, C. M. Cornejo, C. D. Schmults, S. T. Arron, A. Jambusaria-Pahlajani, Field cancerization: Definition, epidemiology, risk factors, and outcomes, J. Am. Acad. Dermatol., 83 (2020), 709–717. https://doi.org/10.1016/j.jaad.2020.03.126 doi: 10.1016/j.jaad.2020.03.126
|
| [4] |
X. Qiu, Z. Gao, J. Shao, H. Li, NCAPH is upregulated in endometrial cancer and associated with poor clinicopathologic characteristics, Ann. Hum. Genet., 84 (2020), 437–446. https://doi.org/10.1111/ahg.12398 doi: 10.1111/ahg.12398
|
| [5] |
Y. Sun, X. Wang, H. Wen, B. Zhu, L. Yu, Expression and clinical significance of the NCAPH, AGGF1, and FOXC2 proteins in serous ovarian cancer, Cancer Manag. Res., 13 (2021), 7253–7262. https://doi.org/10.2147/CMAR.S329688 doi: 10.2147/CMAR.S329688
|
| [6] |
Q. Xiong, S. Fan, L. Duan, B. Liu, X. Jiang, X. Chen, et al., NCAPH is negatively associated with Mcl-1 in non-small cell lung cancer, Mol. Med. Rep., 22 (2020), 2916–2924. https://doi.org/10.3892/mmr.2020.11359 doi: 10.3892/mmr.2020.11359
|
| [7] |
B. Kim, S. W. Kim, J. Y. Lim, S. J. Park, NCAPH is required for proliferation, migration and invasion of non-small-cell lung cancer cells, Anticancer Res., 40 (2020), 3239–3246. https://doi.org/10.21873/anticanres.14305 doi: 10.21873/anticanres.14305
|
| [8] |
M. Wang, X. Qiao, T. Cooper, W. Pan, L. Liu, J. Hayball, et al., HPV E7-mediated NCAPH ectopic expression regulates the carcinogenesis of cervical carcinoma via PI3K/AKT/SGK pathway, Cell Death Dis., 11 (2020), 1049. https://doi.org/10.1038/s41419-020-03244-9 doi: 10.1038/s41419-020-03244-9
|
| [9] |
B. Li, Q. Xiao, L. Shan, Y. Song, NCAPH promotes cell proliferation and inhibits cell apoptosis of bladder cancer cells through MEK/ERK signaling pathway, Cell Cycle, 21 (2022), 427–438. https://doi.org/10.1080/15384101.2021.2021050 doi: 10.1080/15384101.2021.2021050
|
| [10] |
W. Zhou, J. Hu, J. Zhao, Non-SMC condensin I complex subunit H (NCAPH), a regulator of cell cycle, predicts poor prognosis in lung adenocarcinoma patients: a study mainly based on TCGA and GEO database, Transl. Cancer Res., 9 (2020), 7572–7587. https://doi.org/10.21037/tcr-20-2217 doi: 10.21037/tcr-20-2217
|
| [11] |
C. Sun, S. Huang, H. Wang, R. Xie, L. Zhang, Q. Zhou, et al., Non-SMC condensin I complex subunit H enhances proliferation, migration, and invasion of hepatocellular carcinoma, Mol. Carcinog., 58 (2019), 2266–2275. https://doi.org/10.1002/mc.23114 doi: 10.1002/mc.23114
|
| [12] |
Y. Wang, J. Q. Li, Z. L. Yang, L. Wang, J. C. Zhang, Y. F. Sun, et al., NCAPH regulates gastric cancer progression through DNA damage response, Neoplasma, 69 (2021), 283–291. https://doi.org/10.4149/neo_2021_210607N761 doi: 10.4149/neo_2021_210607N761
|
| [13] |
A. Rojas, P. Araya, I. Gonzalez, E. Morales, Gastric tumor microenvironment, Adv. Exp. Med. Biol., 1226 (2020), 23–35. https://doi.org/10.1007/978-3-030-36214-0_2 doi: 10.1007/978-3-030-36214-0_2
|
| [14] |
R. J. DeBerardinis, Tumor microenvironment, metabolism, and immunotherapy, N. Engl. J. Med., 382 (2020), 869–871. https://doi.org/10.1056/NEJMcibr1914890 doi: 10.1056/NEJMcibr1914890
|
| [15] |
Y. Ding, Y. Yan, Y. Dong, J. Xu, W. Su, W. Shi, et al., NLRP3 promotes immune escape by regulating immune checkpoints: A pan-cancer analysis, Int. Immunopharmacol., 104 (2022), 108512. https://doi.org/10.1016/j.intimp.2021.108512 doi: 10.1016/j.intimp.2021.108512
|
| [16] |
X. Nie, M. Zheng, L. Gao, Y. Hu, Y. Zhuang, X. Li, et al., Interaction between TMEFF1 and AHNAK proteins in ovarian cancer cells: Implications for clinical prognosis, Int. Immunopharmacol., 107 (2022), 108726. https://doi.org/10.1016/j.intimp.2022.108726 doi: 10.1016/j.intimp.2022.108726
|
| [17] |
X. F. Wang, W. Lei, C. M. Liu, J. Yang, Y. H. Zhu, BOLA3 is a prognostic-related biomarker and correlated with immune infiltrates in lung adenocarcinoma, Int. Immunopharmacol., 107 (2022), 108652. https://doi.org/10.1016/j.intimp.2022.108652 doi: 10.1016/j.intimp.2022.108652
|
| [18] |
P. Brlek, A. Kafka, A. Bukovac, N. Pećina-Šlaus, Integrative cBioPortal analysis revealed molecular mechanisms that regulate EGFR-PI3K-AKT-mTOR pathway in diffuse gliomas of the brain, Cancers (Basel), 13 (2021), 3247. https://doi.org/10.3390/cancers13133247 doi: 10.3390/cancers13133247
|
| [19] |
J. Watson, J. M. Schwartz, C. Francavilla, Using multilayer heterogeneous networks to infer functions of phosphorylated sites, J. Proteome Res., 20 (2021), 3532–3548. https://doi.org/10.1021/acs.jproteome.1c00150 doi: 10.1021/acs.jproteome.1c00150
|
| [20] |
X. Zhou, J. Du, C. Liu, H. Zeng, Y. Chen, L. Liu, et al., A pan-cancer analysis of CD161, a potential new immune checkpoint, Front. Immunol., 12 (2021), 688215. https://doi.org/10.3389/fimmu.2021.688215 doi: 10.3389/fimmu.2021.688215
|
| [21] |
M. He, Y. Han, C. Cai, P. Liu, Y. Chen, H. Shen, et al., CLEC10A is a prognostic biomarker and correlated with clinical pathologic features and immune infiltrates in lung adenocarcinoma, J. Cell. Mol. Med., 25 (2021), 3391–3399. https://doi.org/10.1111/jcmm.16416 doi: 10.1111/jcmm.16416
|
| [22] |
D. Szklarczyk, A. L. Gable, K. C. Nastou, D. Lyon, R. Kirsch, S. Pyysalo, et al., The STRING database in 2021: customizable protein-protein networks, and functional characterization of user-uploaded gene/measurement sets, Nucleic Acids Res., 49 (2021), D605–D612. https://doi.org/10.1093/nar/gkaa1074 doi: 10.1093/nar/gkaa1074
|
| [23] |
A. Jia, L. Xu, Y. Wang, Venn diagrams in bioinformatics, Brief. Bioinf., 22 (2021), bbab108. https://doi.org/10.1093/bib/bbab108 doi: 10.1093/bib/bbab108
|
| [24] |
W. Liang, F. Sun, Y. Zhao, L. Shan, H. Lou, Identification of susceptibility modules and genes for cardiovascular disease in diabetic patients using WGCNA analysis, J. Diabetes Res., 2020 (2020), 4178639. https://doi.org/10.1155/2020/4178639 doi: 10.1155/2020/4178639
|
| [25] |
C. van Mourik, R. Ehsani, F. Drabløs, GAPGOM—an R package for gene annotation prediction using GO Metrics, BMC Res. Notes, 14 (2021), 162. https://doi.org/10.1186/s13104-021-05580-1 doi: 10.1186/s13104-021-05580-1
|
| [26] |
C. Chen, J. Hou, J. J. Tanner, J. Cheng, Bioinformatics methods for mass spectrometry-based proteomics data analysis, Int. J. Mol. Sci., 21 (2020), 2873. https://doi.org/10.3390/ijms21082873 doi: 10.3390/ijms21082873
|
| [27] |
A. A. Tavakoli, Einstieg ins programmieren für radiologen mit der software R, Die Radiol., 61 (2021), 296–299. https://doi.org/10.1007/s00117-021-00813-7 doi: 10.1007/s00117-021-00813-7
|
| [28] |
Y. Qi, K. Mo, T. Zhang, A transcription factor that promotes proliferation, migration, invasion, and epithelial-mesenchymal transition of ovarian cancer cells and its possible mechanisms, Biomed. Eng. Online, 20 (2021), 83. https://doi.org/10.1186/s12938-021-00919-y doi: 10.1186/s12938-021-00919-y
|
| [29] |
H. Lu, C. Shi, S. Wang, C. Yang, X. Wan, Y. Luo, et al., Identification of NCAPH as a biomarker for prognosis of breast cancer, Mol. Biol. Rep., 47 (2020), 7831–7842. https://doi.org/10.1007/s11033-020-05859-9 doi: 10.1007/s11033-020-05859-9
|
| [30] |
F. Cui, J. Hu, Z. Xu, J. Tan, H. Tang, Overexpression of NCAPH is upregulated and predicts a poor prognosis in prostate cancer, Oncol. Lett., 17 (2019), 5768–5776. https://doi.org/10.3892/ol.2019.10260 doi: 10.3892/ol.2019.10260
|
| [31] |
W. Na, H. Moon, D. Song, A comprehensive review of SARS-CoV-2 genetic mutations and lessons from animal coronavirus recombination in one health perspective, J. Microbiol., 59 (2021), 332–340. https://doi.org/10.1007/s12275-021-0660-4 doi: 10.1007/s12275-021-0660-4
|
| [32] |
L. J. Jilderda, L. Zhou, F. Foijer, Understanding how genetic mutations collaborate with genomic instability in cancer, Cells, 10 (2021), 342. https://doi.org/10.3390/cells10020342 doi: 10.3390/cells10020342
|
| [33] |
C. A. Martin, J. E. Murray, P. Carroll, A. Leitch, K. J. Mackenzie, M. Halachev, Mutations in genes encoding condensin complex proteins cause microcephaly through decatenation failure at mitosis, Genes Dev., 30 (2016), 2158–2172. https://doi.org/10.1101/gad.286351.116 doi: 10.1101/gad.286351.116
|
| [34] |
K. Shimizu, H. Shirataki, T. Honda, S. Minami, Y. Takai, Complex formation of SMAP/KAP3, a KIF3A/B ATPase motor-associated protein, with a human chromosome-associated polypeptide, J. Biol. Chem., 273 (1998), 6591–6594. https://doi.org/10.1074/jbc.273.12.6591 doi: 10.1074/jbc.273.12.6591
|
| [35] |
J. A. Schmiesing, H. C. Gregson, S. Zhou, K. Yokomori, A human condensin complex containing hCAP-C-hCAP-E and CNAP1, a homolog of Xenopus XCAP-D2, colocalizes with phosphorylated histone H3 during the early stage of mitotic chromosome condensation, Mol. Cell. Biol., 20 (2000), 6996–7006. https://doi.org/10.1128/MCB.20.18.6996-7006.2000 doi: 10.1128/MCB.20.18.6996-7006.2000
|
| [36] |
O. A. Cabello, E. Eliseeva, W. G. He, H. Youssoufian, S. E. Plon, B. R. Brinkley, et al., Cell cycle-dependent expression and nucleolar localization of hCAP-H, Mol. Biol. Cell., 12 (2001), 3527–3537. https://doi.org/10.1091/mbc.12.11.3527 doi: 10.1091/mbc.12.11.3527
|
| [37] |
S. H. Chen, W. T. Huang, W. C. Kao, S. Y. Hsiao, H. Y. Pan, C. W. Fang, et al., O6-methylguanine-DNA methyltransferase modulates cisplatin-induced DNA double-strand breaks by targeting the homologous recombination pathway in nasopharyngeal carcinoma, J. Biomed. Sci., 28 (2021), 2. https://doi.org/10.1186/s12929-020-00699-y doi: 10.1186/s12929-020-00699-y
|
| [38] |
X. Huang, Q. Zhang, Y. Lou, J. Wang, X. Zhao, L. Wang, et al., USP22 deubiquitinates CD274 to suppress anticancer immunity, Cancer Immunol. Res., 7 (2019), 1580–1590. https://doi.org/10.1158/2326-6066.CIR-18-0910 doi: 10.1158/2326-6066.CIR-18-0910
|
| [39] |
Q. Liu, T. Gu, L. Y. Su, L. Jiao, X. Qiao, M. Xu, et al., GSNOR facilitates antiviral innate immunity by restricting TBK1 cysteine S-nitrosation, Redox Biol., 47 (2021), 102172. https://doi.org/10.1016/j.redox.2021.102172 doi: 10.1016/j.redox.2021.102172
|
| [40] |
M. Z. Jin, Y. G. Zhang, W. L. Jin, X. P. Wang, A pan-cancer analysis of the oncogenic and immunogenic role of m6Am methyltransferase PCIF1, Front. Oncol., 11 (2021), 753393. https://doi.org/10.3389/fonc.2021.753393 doi: 10.3389/fonc.2021.753393
|
| [41] |
L. Yin, L. P. Jiang, Q. S. Shen, Q. X. Xiong, X. Zhuo, L. L. Zhang, et al., NCAPH plays important roles in human colon cancer, Cell Death Dis., 8 (2017), e2680. https://doi.org/10.1038/cddis.2017.88 doi: 10.1038/cddis.2017.88
|
| [42] |
M. Casanova-Acebes, E. Dalla, A. M. Leader, J. LeBerichel, J. Nikolic, B. M. Morales, et al., Tissue-resident macrophages provide a pro-tumorigenic niche to early NSCLC cells, Nature, 595 (2021), 578–584. https://doi.org/10.1038/s41586-021-03651-8 doi: 10.1038/s41586-021-03651-8
|
 mbe-20-01-005-supplementary.pdf mbe-20-01-005-supplementary.pdf |
 |






Figures(7)

Ying Liu, Xiao Ma, Linyuan Feng, Zhenhua Lin, Xianchun Zhou. An integrative pan-cancer analysis reveals the carcinogenic effects of NCAPH in human cancer[J]. Mathematical Biosciences and Engineering, 2023, 20(1): 76-92. doi: 10.3934/mbe.2023005







 DownLoad:
DownLoad: