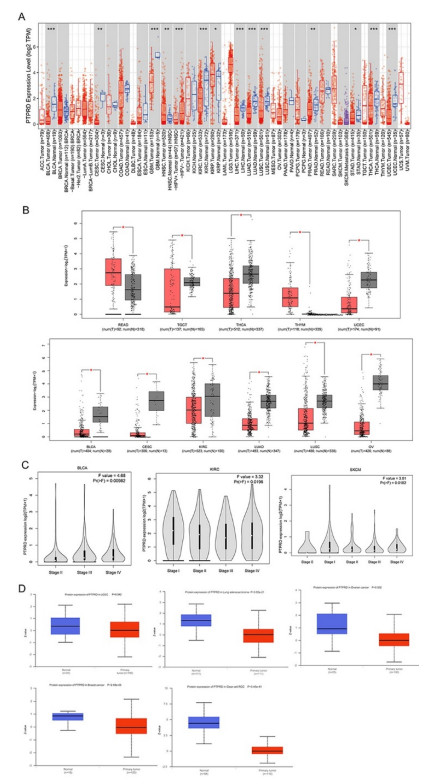
PTPRD plays an indispensable role in the occurrence of multiple tumors. However, pan-cancer analysis is unavailable. The purpose of this research was to preliminarily study its prognostic landscape across various tumors and investigate its relationship with immunotherapy. We exhibited the expression profile, survival analysis, and genomic alterations of PTPRD based on the TIMER, GEPIA, UALCAN, PrognoScan and cBioPortal database. The frequency of PTPRD mutation and its correlation with response to immunotherapy were evaluated using the cBioPortal database. The relationship between PTPRD and immune-cell infiltration was analyzed by the TIMER and TISIDB databases. A protein interaction network was constructed by the STRING database. GO and KEGG enrichment analysis was executed by the Metascape database. A correlation between PTPRD expression and prognosis was found in various cancers. Aberrant PTPRD expression was closely related to immune infiltration. In non-small cell lung cancer and melanoma, patients with PTPRD mutations had better overall survival with immune checkpoint inhibitors, and these patients had higher TMB scores. PTPRD mutation was involved in numerous biological processes, including immunological signaling pathways. A PTPRD protein interaction network was constructed, and genes that interacted with PTPRD were identified. Functional enrichment analysis demonstrated that a variety of GO biological processes and KEGG pathways associated with PTPRD were involved in the therapeutic mechanisms. These results revealed that PTPRD might function as a biomarker for prognosis and immune infiltration in cancers, throwing new light on cancer therapeutics.
Citation: Chunpei Ou, Qin Peng, Changchun Zeng. An integrative prognostic and immune analysis of PTPRD in cancer[J]. Mathematical Biosciences and Engineering, 2022, 19(6): 5361-5379. doi: 10.3934/mbe.2022251
PTPRD plays an indispensable role in the occurrence of multiple tumors. However, pan-cancer analysis is unavailable. The purpose of this research was to preliminarily study its prognostic landscape across various tumors and investigate its relationship with immunotherapy. We exhibited the expression profile, survival analysis, and genomic alterations of PTPRD based on the TIMER, GEPIA, UALCAN, PrognoScan and cBioPortal database. The frequency of PTPRD mutation and its correlation with response to immunotherapy were evaluated using the cBioPortal database. The relationship between PTPRD and immune-cell infiltration was analyzed by the TIMER and TISIDB databases. A protein interaction network was constructed by the STRING database. GO and KEGG enrichment analysis was executed by the Metascape database. A correlation between PTPRD expression and prognosis was found in various cancers. Aberrant PTPRD expression was closely related to immune infiltration. In non-small cell lung cancer and melanoma, patients with PTPRD mutations had better overall survival with immune checkpoint inhibitors, and these patients had higher TMB scores. PTPRD mutation was involved in numerous biological processes, including immunological signaling pathways. A PTPRD protein interaction network was constructed, and genes that interacted with PTPRD were identified. Functional enrichment analysis demonstrated that a variety of GO biological processes and KEGG pathways associated with PTPRD were involved in the therapeutic mechanisms. These results revealed that PTPRD might function as a biomarker for prognosis and immune infiltration in cancers, throwing new light on cancer therapeutics.

| [1] |
R. L. Siegel, K. D. Miller, H. E. Fuchs, A. Jemal, Cancer Statistics, 2021, CA Cancer J. Clin., 71 (2021), 7-33. https://doi.org/10.3322/caac.21654 doi: 10.3322/caac.21654

|
| [2] |
P. Krzyszczyk, A. Acevedo, E. J. Davidoff, L. M. Timmins, I. Marrero-Berrios, M. Patel, et al., The growing role of precision and personalized medicine for cancer treatment, Technology, 6 (2018), 79-100. https://doi.org/10.1142/S2339547818300020 doi: 10.1142/S2339547818300020
|
| [3] |
The Cancer Genome Atlas Research Network, J. N. Weinstein, E. A. Collisson, G. B. Mills, K. R. Shaw, B. A. Ozenberger, et al., The cancer genome atlas pan-cancer analysis project, Nat. Genet., 45 (2013), 1113-1120. https://doi.org/10.1038/ng.2764 doi: 10.1038/ng.2764
|
| [4] |
J. Liu, T. Lichtenberg, K. A. Hoadley, L. M. Poisson, A. J. Lazar, A. D. Cherniack, et al., An integrated TCGA pan-cancer clinical data resource to drive high-quality survival outcome analytics, Cell, 173 (2018), 400-416. https://doi.org/10.1158/1538-7445.AM2018-3287 doi: 10.1158/1538-7445.AM2018-3287
|
| [5] |
F. Chen, M. C. Wendl, M. A. Wyczalkowski, M. H. Bailey, Y. Li, L. Ding, Moving pan-cancer studies from basic research toward the clinic, Nat. Cancer, 2 (2021), 879-890. https://doi.org/10.1038/s43018-021-00250-4 doi: 10.1038/s43018-021-00250-4
|
| [6] |
S. Veeriah, C. Brennan, S. Meng, B. Singh, J. A. Fagin, D. B. Solit, et al., The tyrosine phosphatase PTPRD is a tumor suppressor that is frequently inactivated and mutated in glioblastoma and other human cancers, Proc. Natl. Acad. Sci., 106 (2009), 9435-9440. https://doi.org/10.1073/pnas.0900571106 doi: 10.1073/pnas.0900571106
|
| [7] |
X. Huang, F. Qin, Q. Meng, M. Dong, Protein tyrosine phosphatase receptor type D (PTPRD)-mediated signaling pathways for the potential treatment of hepatocellular carcinoma: a narrative review, Ann. Transl. Med., 8 (2020), 1192. https://doi.org/10.21037/atm-20-4733 doi: 10.21037/atm-20-4733
|
| [8] |
G. R. Uhl, M. J. Martinez, PTPRD: neurobiology, genetics, and initial pharmacology of a pleiotropic contributor to brain phenotypes, Ann. N. Y. Acad. Sci., 1451 (2019), 112-129. https://doi.org/10.1111/nyas.14002 doi: 10.1111/nyas.14002
|
| [9] |
K. Funato, Y. Yamazumi, T. Oda, T. Akiyama, Tyrosine phosphatase PTPRD suppresses colon cancer cell migration in coordination with CD44, Exp. Ther. Med., 2 (2011), 457-463. https://doi.org/10.3892/etm.2011.231 doi: 10.3892/etm.2011.231
|
| [10] |
W. J. Bae, J. M. Ahn, H. E. Byeon, S. Kim, D. Lee, PTPRD-inactivation-induced CXCL8 promotes angiogenesis and metastasis in gastric cancer and is inhibited by metformin, J. Exp. Clin. Cancer Res., 38 (2019), 484. https://doi.org/10.1186/s13046-019-1469-4 doi: 10.1186/s13046-019-1469-4
|
| [11] |
K. Onoi, Y. Chihara, J. Uchino, T. Shimamoto, Y. Morimoto, M. Iwasaku, et al., Immune checkpoint inhibitors for lung cancer treatment: A review, J. Clin. Med., 9 (2020). https://doi.org/10.3390/jcm9051362 doi: 10.3390/jcm9051362
|
| [12] |
A. M. Goodman, E. S. Sokol, G. M. Frampton, S. M. Lippman, R. Kurzrock, Microsatellite-stable tumors with high mutational burden benefit from immunotherapy, Cancer Immunol. Res., 7 (2019), 1570-1573. https://doi.org/10.1158/2326-6066.CIR-19-0149 doi: 10.1158/2326-6066.CIR-19-0149
|
| [13] |
D. Sha, Z. Jin, J. Budczies, K. Kluck, A. Stenzinger, F. A. Sinicrope, Tumor mutational burden as a predictive biomarker in solid tumors, Cancer Discov., 10 (2020), 1808-1825. https://doi.org/10.1158/2159-8290.CD-20-0522 doi: 10.1158/2159-8290.CD-20-0522
|
| [14] |
Z. Lu, H. Chen, X. Jiao, W. Zhou, W. Han, S. Li, et al., Prediction of immune checkpoint inhibition with immune oncology-related gene expression in gastrointestinal cancer using a machine learning classifier, J. Immunother. Cancer, 8 (2020). https://doi.org/10.1136/jitc-2020-000631 doi: 10.1136/jitc-2020-000631
|
| [15] |
N. Ready, M. D. Hellmann, M. M. Awad, G. A. Otterson, M. Gutierrez, J. F. Gainor, et al., First-line nivolumab plus ipilimumab in advanced non-small-cell lung cancer (CheckMate 568): outcomes by programmed death ligand 1 and tumor mutational burden as biomarkers, J. Clin. Oncol., 37 (2019), 992-1000. https://doi.org/10.1200/JCO.18.01042 doi: 10.1200/JCO.18.01042
|
| [16] |
D. A. Solomon, J. S. Kim, J. C. Cronin, Z. Sibenaller, T. Ryken, S. A. Rosenberg, et al., Mutational inactivation of PTPRD in glioblastoma multiforme and malignant melanoma, Cancer Res., 68 (2008), 10300-10306. https://doi.org/10.1158/0008-5472.CAN-08-3272 doi: 10.1158/0008-5472.CAN-08-3272
|
| [17] |
T. Li, J. Fu, Z. Zeng, D. Cohen, J. Li, Q. Chen, et al., TIMER2.0 for analysis of tumor-infiltrating immune cells, Nucleic Acids Res., 48 (2020), W509-W514. https://doi.org/10.1093/nar/gkaa407 doi: 10.1093/nar/gkaa407
|
| [18] |
D. S. Chandrashekar, B. Bashel, S. A. H. Balasubramanya, C. J. Creighton, I. Ponce-Rodriguez, B. Chakravarthi, et al., UALCAN: a portal for facilitating tumor subgroup gene expression and survival analyses, Neoplasia, 19 (2017), 649-658. https://doi.org/10.1016/j.neo.2017.05.002 doi: 10.1016/j.neo.2017.05.002
|
| [19] |
H. Mizuno, K. Kitada, K. Nakai, A. Sarai, PrognoScan: a new database for meta-analysis of the prognostic value of genes, BMC Med. Genomics, 2 (2009), 18. https://doi.org/10.1186/1755-8794-2-18 doi: 10.1186/1755-8794-2-18
|
| [20] |
J. Gao, B. A. Aksoy, U. Dogrusoz, G. Dresdner, B. Gross, S. O. Sumer, et al., Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal, Sci. Signal, 6 (2013). https://doi.org/10.1126/scisignal.2004088 doi: 10.1126/scisignal.2004088
|
| [21] |
R. M. Samstein, C. H. Lee, A. N. Shoushtari, M. D. Hellmann, R. Shen, Y. Y. Janjigian, et al., Tumor mutational load predicts survival after immunotherapy across multiple cancer types, Nat. Genet., 51 (2019), 202-206. https://doi.org/10.1038/s41588-018-0312-8 doi: 10.1038/s41588-018-0312-8
|
| [22] |
D. Miao, C. A. Margolis, N. I. Vokes, D. Liu, A. Taylor-Weiner, S. M. Wankowicz, et al., Genomic correlates of response to immune checkpoint blockade in microsatellite-stable solid tumors, Nat. Genet., 50 (2018), 1271-1281. https://doi.org/10.1038/s41588-018-0200-2 doi: 10.1038/s41588-018-0200-2
|
| [23] |
H. Rizvi, F. Sanchez-Vega, K. La, W. Chatila, P. Jonsson, D. Halpenny, et al., Molecular determinants of response to anti-programmed cell death (PD)-1 and anti-programmed death-ligand 1 (PD-L1) blockade in patients with non-small-cell lung cancer profiled with targeted next-generation sequencing, J. Clin. Oncol., 36 (2018), 633-641. https://doi.org/10.1200/JCO.2017.75.3384 doi: 10.1200/JCO.2017.75.3384
|
| [24] |
N. A. Rizvi, M. D. Hellmann, A. Snyder, P. Kvistborg, V. Makarov, J. J. Havel, et al., Mutational landscape determines sensitivity to PD-1 blockade in non-small cell lung cancer, Science, 348 (2015), 124-128. https://doi.org/10.1126/science.aaa1348 doi: 10.1126/science.aaa1348
|
| [25] |
T. Li, J. Fan, B. Wang, N. Traugh, Q. Chen, J. S. Liu, et al., TIMER: a web server for comprehensive analysis of tumor-infiltrating immune cells, Cancer Res., 77 (2017), e108-e110. https://doi.org/10.1158/0008-5472.CAN-17-0307 doi: 10.1158/0008-5472.CAN-17-0307
|
| [26] |
B. Li, E. Severson, J. C. Pignon, H. Zhao, T. Li, J. Novak, et al., Comprehensive analyses of tumor immunity: implications for cancer immunotherapy, Genome Biol., 17 (2016), 174. https://doi.org/10.1186/s13059-016-1028-7 doi: 10.1186/s13059-016-1028-7
|
| [27] |
B. Ru, C. N. Wong, Y. Tong, J. Y. Zhong, S. S. W. Zhong, W. C. Wu, et al., TISIDB: an integrated repository portal for tumor-immune system interactions, Bioinformatics, 35 (2019), 4200-4202. https://doi.org/10.1093/bioinformatics/btz210 doi: 10.1093/bioinformatics/btz210
|
| [28] |
Z. Tang, C. Li, B. Kang, G. Gao, C. Li, Z. Zhang, GEPIA: a web server for cancer and normal gene expression profiling and interactive analyses, Nucleic Acids Res., 45 (2017), W98-W102. https://doi.org/10.1093/nar/gkx247 doi: 10.1093/nar/gkx247
|
| [29] |
D. Szklarczyk, A. L. Gable, D. Lyon, A. Junge, S. Wyder, J. Huerta-Cepas, et al., STRING v11: protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets, Nucleic Acids Res., 47 (2019), D607-D613. https://doi.org/10.1093/nar/gky1131 doi: 10.1093/nar/gky1131
|
| [30] |
Y. Zhou, B. Zhou, L. Pache, M. Chang, A. H. Khodabakhshi, O. Tanaseichuk, et al., Metascape provides a biologist-oriented resource for the analysis of systems-level datasets, Nat. Commun., 10 (2019), 1523. https://doi.org/10.1038/s41467-019-09234-6 doi: 10.1038/s41467-019-09234-6
|
| [31] |
T. Acun, K. Demir, E. Oztas, D. Arango, M. C. Yakicier, PTPRD is homozygously deleted and epigenetically downregulated in human hepatocellular carcinomas, OMICS, 19 (2015), 220-229. https://doi.org/10.1089/omi.2015.0010 doi: 10.1089/omi.2015.0010
|
| [32] |
H. Tomita, F. Cornejo, B. Aranda-Pino, C. L. Woodard, C. C. Rioseco, B. G. Neel, et al., The protein tyrosine phosphatase receptor delta regulates developmental neurogenesis, Cell Rep., 30 (2020), 215-228. https://doi.org/10.1016/j.celrep.2019.11.033 doi: 10.1016/j.celrep.2019.11.033
|
| [33] |
H. C. Hsu, N. Lapke, S. J. Chen, Y. J. Lu, R. S. Jhou, C. Y. Yeh, et al., PTPRT and PTPRD deleterious mutations and deletion predict bevacizumab resistance in metastatic colorectal cancer patients, Cancers (Basel), 10 (2018). https://doi.org/10.3390/cancers10090314 doi: 10.3390/cancers10090314
|
| [34] |
G. R. Uhl, M. J. Martinez, P. Paik, A. Sulima, G. H. Bi, M. R. Iyer, et al., Cocaine reward is reduced by decreased expression of receptor-type protein tyrosine phosphatase D (PTPRD) and by a novel PTPRD antagonist, Proc. Natl. Acad. Sci., 115 (2018), 11597-11602. https://doi.org/10.1073/pnas.1720446115 doi: 10.1073/pnas.1720446115
|
| [35] |
N. D. Peyser, Y. Du, H. Li, V. Lui, X. Xiao, T. A. Chan, et al., Loss-of-function PTPRD mutations lead to increased STAT3 activation and sensitivity to STAT3 inhibition in head and neck cancer, PLoS One, 10 (2015), e0135750. https://doi.org/10.1371/journal.pone.0135750 doi: 10.1371/journal.pone.0135750
|
| [36] |
L. Wu, L. Gao, D. Kong, H. Xue, Loss of tyrosine phosphatase delta promotes gastric cancer progression via signal transducer and activator of transcription 3 pathways, Dig. Dis. Sci., 64 (2019), 3164-3172. https://doi.org/10.1007/s10620-019-05637-z doi: 10.1007/s10620-019-05637-z
|
| [37] |
B. Ortiz, A. W. Fabius, W. H. Wu, A. Pedraza, C. W. Brennan, N. Schultz, et al., Loss of the tyrosine phosphatase PTPRD leads to aberrant STAT3 activation and promotes gliomagenesis, Proc. Natl. Acad. Sci., 111 (2014), 8149-8154. https://doi.org/10.1073/pnas.1401952111 doi: 10.1073/pnas.1401952111
|
| [38] |
F. Zhang, B. Wang, T. Qin, L. Wang, Q. Zhang, Y. Lu, et al., IL-6 induces tumor suppressor protein tyrosine phosphatase receptor type D by inhibiting miR-34a to prevent IL-6 signaling overactivation, Mol. Cell Biochem., 473 (2020). https://doi.org/10.1007/s11010-020-03803-w doi: 10.1007/s11010-020-03803-w
|
| [39] |
L. Ding, X. Chen, X. Xu, Y. Qian, G. Liang, F. Yao, et al., PARP1 suppresses the transcription of PD-L1 by poly(ADP-Ribosyl) ating STAT3, Cancer Immunol. Res., 7 (2019), 136-149. https://doi.org/10.1158/2326-6066.CIR-18-0071 doi: 10.1158/2326-6066.CIR-18-0071
|
| [40] |
J. Mo, X. Hu, L. Gu, B. Chen, P. A. Khadaroo, Z. Shen, et al., Smokers or non-smokers: who benefits more from immune checkpoint inhibitors in treatment of malignancies? An up-to-date meta-analysis, World J. Surg. Oncol., 18 (2020), 15. https://doi.org/10.1186/s12957-020-1792-4 doi: 10.1186/s12957-020-1792-4
|
| [41] |
G. T. Gibney, L. M. Weiner, M. B. Atkins, Predictive biomarkers for checkpoint inhibitor-based immunotherapy, Lancet Oncol., 17 (2016), e542-e551. https://doi.org/10.1016/S1470-2045(16)30406-5 doi: 10.1016/S1470-2045(16)30406-5
|
| [42] |
Y. Qiu, L. Liu, H. Yang, H. Chen, Q. Deng, D. Xiao, et al., Integrating histologic and genomic characteristics to predict tumor mutation burden of early-stage non-small-cell lung cancer, Front. Oncol., 10 (2020), 608989. https://doi.org/10.3389/fonc.2020.608989 doi: 10.3389/fonc.2020.608989
|
| [43] |
J. Norum, C. Nieder, Tobacco smoking and cessation and PD-L1 inhibitors in non-small cell lung cancer (NSCLC): a review of the literature, ESMO Open, 3 (2018), e000406. https://doi.org/10.1136/esmoopen-2018-000406 doi: 10.1136/esmoopen-2018-000406
|
 mbe-19-06-251-supplementary.pdf mbe-19-06-251-supplementary.pdf |
 |






Figures(7)

Chunpei Ou, Qin Peng, Changchun Zeng. An integrative prognostic and immune analysis of PTPRD in cancer[J]. Mathematical Biosciences and Engineering, 2022, 19(6): 5361-5379. doi: 10.3934/mbe.2022251







 DownLoad:
DownLoad: