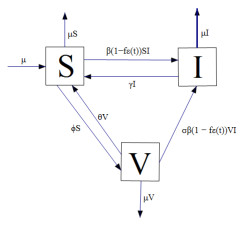
This study aims to analyze a class of SIV systems considering the transmission rate influenced by media coverage and protective measures, in which the transmission rate is represented by a piecewise-smooth function. Firstly, for the SIV Filippov system, we take the dynamic behaviors of two subsystems into consideration, and obtain the basic reproduction number and the equilibria of the subsystems respectively. Secondly, based on the Filippov convex method, we calculate the sliding domain and the sliding mode equation, and further analyze the global dynamic behaviors of the system, through which we verify that there is no closed orbit in the system. Furthermore, we prove the global asymptotical stability of the disease-free equilibrium, two real equilibria, and the pseudo-equilibrium under certain conditions. The results demonstrate that the threshold value, the protective measures, and the media coverage could affect the number of infected individuals and the final scale of the disease. To prevent the spread of the disease, it is necessary to select an appropriate threshold and take applicable protective measures combined with media coverage. Lastly, we verify the validity of the results by numerical simulations.
Citation: Shifan Luo, Dongshu Wang, Wenxiu Li. Dynamic analysis of a SIV Filippov system with media coverage and protective measures[J]. AIMS Mathematics, 2022, 7(7): 13469-13492. doi: 10.3934/math.2022745
This study aims to analyze a class of SIV systems considering the transmission rate influenced by media coverage and protective measures, in which the transmission rate is represented by a piecewise-smooth function. Firstly, for the SIV Filippov system, we take the dynamic behaviors of two subsystems into consideration, and obtain the basic reproduction number and the equilibria of the subsystems respectively. Secondly, based on the Filippov convex method, we calculate the sliding domain and the sliding mode equation, and further analyze the global dynamic behaviors of the system, through which we verify that there is no closed orbit in the system. Furthermore, we prove the global asymptotical stability of the disease-free equilibrium, two real equilibria, and the pseudo-equilibrium under certain conditions. The results demonstrate that the threshold value, the protective measures, and the media coverage could affect the number of infected individuals and the final scale of the disease. To prevent the spread of the disease, it is necessary to select an appropriate threshold and take applicable protective measures combined with media coverage. Lastly, we verify the validity of the results by numerical simulations.

| [1] |
G. Zaman, Y. H. Kang, I. H. Jung, Stability analysis and optimal vaccination of an SIR epidemic model, Biosystems, 93 (2008), 240â€"249. http://dx.doi.org/10.1016/j.biosystems.2008.05.004 doi: 10.1016/j.biosystems.2008.05.004

|
| [2] |
I. Cooper, A. Mondal, C. G. Antonopoulos, A SIR model assumption for the spread of COVID-19 in different communities, Chaos Soliton. Fract., 139 (2020), 110057. http://dx.doi.org/10.1016/j.chaos.2020.110057 doi: 10.1016/j.chaos.2020.110057
|
| [3] |
L. J. S Allen, Some discrete-time SI, SIR, and SIS epidemic models, Math. Biosci., 124 (1994), 83â€"105. http://dx.doi.org/10.1016/0025-5564(94)90025-6 doi: 10.1016/0025-5564(94)90025-6
|
| [4] |
Y. Enatsu, Y. Nakata, Y. Muroya, Global stability for a class of discrete SIR epidemic models, Math. Biosci. Eng., 7 (2010), 347â€"361. http://dx.doi.org/10.3934/mbe.2010.7.347 doi: 10.3934/mbe.2010.7.347
|
| [5] |
B. Shulgin, L. Stone, Z. Agur, Pulse vaccination strategy in the SIR epidemic model, Bull. Math. Biol., 60 (1998), 1123â€"1148. http://dx.doi.org/10.1006/s0092-8240(98)90005-2 doi: 10.1006/s0092-8240(98)90005-2
|
| [6] |
L. Stone, B. Shulgin, Z. Agur, Theoretical examination of the pulse vaccination policy in the SIR epidemic model, Math. Comput. Model., 31 (2000), 207â€"215. http://dx.doi.org/10.1016/S0895-7177(00)00040-6 doi: 10.1016/S0895-7177(00)00040-6
|
| [7] |
T. Zhao, Y. Xiao, Non-smooth plant disease models with economic thresholds, Math. Biosci., 241 (2013), 34â€"48. http://dx.doi.org/10.1016/j.mbs.2012.09.005 doi: 10.1016/j.mbs.2012.09.005
|
| [8] |
Z. Guo, L. Huang, X. Zou, Impact of discontinuous treatments on disease dynamics in an SIR epidemic model, Math. Biosci. Eng., 9 (2012), 97â€"110. http://dx.doi.org/10.3934/mbe.2012.9.97 doi: 10.3934/mbe.2012.9.97
|
| [9] |
Z. Guo, X. Zou, Impact of discontinuous harvesting on fishery dynamics in a stock-effort fishing model, Commun. Nonlinear Sci., 20 (2015), 594â€"603. http://dx.doi.org/10.1016/j.cnsns.2014.06.014 doi: 10.1016/j.cnsns.2014.06.014
|
| [10] |
L. Huang, H. Ma, J. Wang, C. Huang, Global dynamics of a Filippov plant disease model with an economic threshold of infected-susceptible ratio, J. Appl. Anal. Comput., 10 (2020), 2263â€"2277. http://dx.doi.org/10.11948/20190409 doi: 10.11948/20190409
|
| [11] |
W. Li, L. Huang, J. Wang, Global dynamics of Filippov-type plant disease models with an interaction ratio threshold, Math. Meth. Appl. Sci., 43 (2020), 6995â€"7008. http://dx.doi.org/10.1002/mma.6450 doi: 10.1002/mma.6450
|
| [12] |
Y. K. Xie, Z. Wang, A ratio-dependent impulsive control of an SIQS epidemic model with non-linear incidence, Appl. Math. Comput., 423 (2022), 127018. https://doi.org/10.1016/j.amc.2022.127018 doi: 10.1016/j.amc.2022.127018
|
| [13] |
Y. K. Xie, Z. Wang, Transmission dynamics, global stability and control strategies of a modified SIS epidemic model on complex networks with an infective medium, Math. Comput. Simul., 188 (2021), 23â€"34. https://doi.org/10.1016/j.matcom.2021.03.029 doi: 10.1016/j.matcom.2021.03.029
|
| [14] |
W. O. Kermack, A. G. McKendrick, A contribution to the mathematical theory of epidemics, P. Roy. Soc. A, 115 (1927), 700â€"721. http://dx.doi.org/10.1007/bf02464423 doi: 10.1007/bf02464423
|
| [15] |
W. O. Kermack, A. G. McKendrick, Contributions to the mathematical theory of epidemics â…¡.â€"-The problem of endemicity, P. Roy. Soc. A, 138 (1932), 55â€"83. http://dx.doi.org/10.1016/s0092-8240(05)80041-2 doi: 10.1016/s0092-8240(05)80041-2
|
| [16] |
W. O. Kermack, A. G. McKendrick, Contributions to the mathematical theory of epidemics â…¢.â€"-Further studies of the problem of endemicity, P. Roy. Soc. A, 141 (1933), 94â€"122. http://dx.doi.org/10.2307/96207 doi: 10.2307/96207
|
| [17] |
B Shulgin, L. W. Stone, Z. Agur, Pulse vaccination strategy in the SIR epidemic model, Bull. Math. Biol., 60 (1998), 1123â€"1148. http://dx.doi.org/10.1016/S0092-8240(98)90005-2 doi: 10.1016/S0092-8240(98)90005-2
|
| [18] |
A. B. Gumel, S. M. Moghadas, A qualitative study of a vaccination model with non-linear incidence, Appl. Math. Comput., 143 (2003), 409â€"419. http://dx.doi.org/10.1016/S0096-3003(02)00372-7 doi: 10.1016/S0096-3003(02)00372-7
|
| [19] |
S. Tang, J. Liang, Y. Xiao, R. A. Cheke, Sliding bifurcations of Filippov two stage pest control models with economic thresholds, SIAM J. Appl. Math., 72 (2012), 1061â€"1080. http://dx.doi.org/10.1137/110847020 doi: 10.1137/110847020
|
| [20] |
Y. Zhang, Y. Xiao, Global dynamics for a Filippov epidemic system with imperfect vaccination, Nonlinear Anal. Hybrid Syst., 38 (2020), 100932. http://dx.doi.org/10.1016/j.nahs.2020.100932 doi: 10.1016/j.nahs.2020.100932
|
| [21] |
A. P. Lemos-Paio, C. J. Silva, D. F. M. Torres, An epidemic model for cholera with optimal control treatment, J. Comput. Appl. Math., 318 (2017), 168â€"180. http://dx.doi.org/10.1016/j.cam.2016.11.002 doi: 10.1016/j.cam.2016.11.002
|
| [22] |
M. J. Jeger, L. V. Madden, F. V. D. Bosch, The effect of transmission route on plant virus epidemic development and disease control, J. Theor. Biol., 258 (2009), 198â€"207. http://dx.doi.org/10.1016/j.jtbi.2009.01.012 doi: 10.1016/j.jtbi.2009.01.012
|
| [23] |
V. C. C. Cheng, S. C. Wong, V. W. M. Chuang, S. Y. C. So, J. H. K. Chen, S. Sridhar, et al., The role of community-wide wearing of face mask for control of coronavirus disease 2019 (COVID-19) epidemic due to SARS-CoV-2, J. Infect., 81 (2020), 107â€"114. http://dx.doi.org/10.1016/j.jinf.2020.04.024 doi: 10.1016/j.jinf.2020.04.024
|
| [24] |
S. E. Eikenberry, M. Mancuso, E. Iboi, T. Phan, K. Eikenberry, Y. Kuang, et al., To mask or not to mask: Modeling the potential for face mask use by the general public to curtail the covid-19 pandemic, Infect. Dis. Model., 5 (2020), 293â€"308. http://dx.doi.org/10.1101/2020.04.06.20055624 doi: 10.1101/2020.04.06.20055624
|
| [25] |
C. R. MacIntyre, S. Cauchemez, D. E. Dwyer, H. Seale, P. Cheung, G. Browne, et al., Face mask use and control of respiratory virus transmission in households, Emerg. Infect. Dis., 15 (2009), 233. http://dx.doi.org/10.3201/eid1502.081167 doi: 10.3201/eid1502.081167
|
| [26] |
Z. Tai, T. Sun, Media dependencies in a changing media environment: The case of the 2003 SARS epidemic in China, New Media Soc., 9 (2007), 987â€"1009. http://dx.doi.org/10.1177/1461444807082691 doi: 10.1177/1461444807082691
|
| [27] |
S. Collinson, J. M. Heffernan, Modelling the effects of media during an influenza epidemic, BMC Public Health, 14 (2014), 1â€"10. http://dx.doi.org/10.1186/1471-2458-14-376 doi: 10.1186/1471-2458-14-376
|
| [28] | R. M. Anderson, R. M. May, Infectious diseases of humans: Dynamics and control, Oxford: Oxford Science Publications, 1992. http://dx.doi.org/10.1126/science.254.5031.591 |
| [29] |
J. Cui, Y. Sun, H. Zhu, The impact of media on the spreading and control of infectious disease, J. Dyn. Differ. Equ., 20 (2008), 31â€"53. http://dx.doi.org/10.1007/s10884-007-9075-0 doi: 10.1007/s10884-007-9075-0
|
| [30] |
J. Deng, S. Tang, H. Shu, Joint impacts of media, vaccination and treatment on an epidemic Filippov model with application to COVID-19, J. Theor. Biol., 523 (2021), 110698. http://dx.doi.org/10.1016/j.jtbi.2021.110698 doi: 10.1016/j.jtbi.2021.110698
|
| [31] |
A. Wang, Y. Xiao, A Filippov system describing media effects on the spread of infectious diseases, Nonlinear Anal. Hybrid Syst., 11 (2014), 84â€"97. http://dx.doi.org/10.1016/j.nahs.2013.06.005 doi: 10.1016/j.nahs.2013.06.005
|
| [32] |
J. Wang, F. Zhang, L. Wang, Equilibrium, pseudoequilibrium and sliding-mode heteroclinic orbit in a Filippov-type plant disease model, Nonlinear Anal.-Real, 31 (2016), 308â€"324. http://dx.doi.org/10.1016/j.nonrwa.2016.01.017 doi: 10.1016/j.nonrwa.2016.01.017
|
| [33] |
W. Li, L. Huang, J. Wang, Dynamic analysis of discontinuous plant disease models with a non-smooth separation line, Nonlinear Dyn., 99 (2020), 1675â€"1697. http://dx.doi.org/10.1007/s11071-019-05384-w doi: 10.1007/s11071-019-05384-w
|
| [34] | A. F. Filippov, Differential equations with discontinuous right-hand side, Dordrecht: Kluwer Academic, 1988. http://dx.doi.org/10.1016/0022-247X(91)90044-Z |
| [35] |
X. Chen, L. Huang, A Filippov system describing the effect of prey refuge use on a ratio-dependent predator-prey model, J. Math. Anal. Appl., 428 (2015), 817â€"837. http://dx.doi.org/10.1016/j.jmaa.2015.03.045 doi: 10.1016/j.jmaa.2015.03.045
|
| [36] |
A. Wang, Y. Xiao, Sliding bifurcation and global dynamics of a Filippov epidemic model with vaccination, Int. J. Bifurcat. Chaos, 23 (2013), 1350144. http://dx.doi.org/10.1142/S0218127413501447 doi: 10.1142/S0218127413501447
|
| [37] |
M. di Bernardo, C. J. Budd, A. R. Champneys, P. Kowalczyk, A. B. Nordmark, Bifurcation in nonsmooth dynamical systems, SIAM Rev., 50 (2008), 629â€"701. http://dx.doi.org/10.1137/050625060 doi: 10.1137/050625060
|
| [38] |
Y. A. Kuznetsov, S. Rinaldi, A. Gragnani, One parameter bifurcations in planar Filippov systems, Int. J. Bifurcat. Chaos, 13 (2003), 2157â€"2188. http://dx.doi.org/10.1142/S0218127403007874 doi: 10.1142/S0218127403007874
|
| [39] |
P. V. D. Driessche, J. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180 (2002), 29â€"48. http://dx.doi.org/10.1016/S0025-5564(02)00108-6 doi: 10.1016/S0025-5564(02)00108-6
|
| [40] | Z. Ma, Y. Zhou, C. Li, Qualitative and stability methods for ordinary differential equations, Beijing: Science Press, 2015. |
| [41] |
R. M. Corless, G. H. Gonnet, D. E. G. Hare, D. J. Jeffrey, D. E. Kunth, On the Lambert W function, Adv. Comput. Math., 5 (1996), 329â€"359. http://dx.doi.org/10.1007/BF02124750 doi: 10.1007/BF02124750
|









Figures(10) / Tables(2)

Shifan Luo, Dongshu Wang, Wenxiu Li. Dynamic analysis of a SIV Filippov system with media coverage and protective measures[J]. AIMS Mathematics, 2022, 7(7): 13469-13492. doi: 10.3934/math.2022745







 DownLoad:
DownLoad: