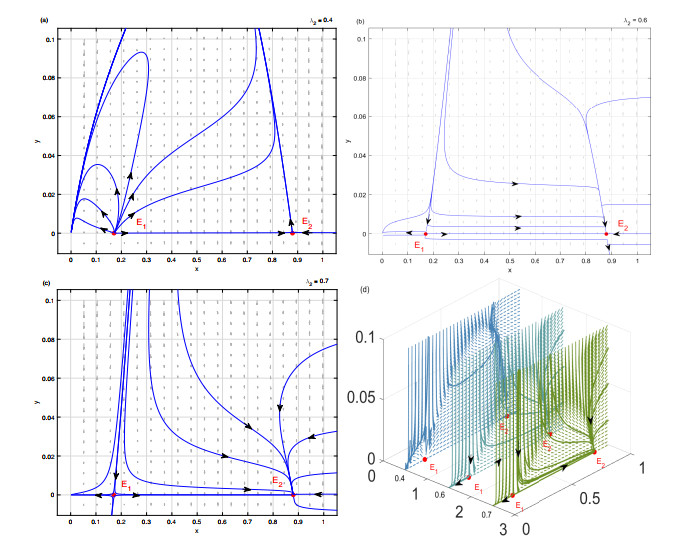
In the paper, a predator-prey model with the Allee effect and harvesting effort was proposed to explore the interaction mechanism between prey and predator. Under the framework of mathematical theory deduction, some conditions for the occurrence of transcritical, saddle-node, Hopf, and Bogdanov-Takens bifurcations were derived with harvesting effort and the Allee effect as key parameters. Under the framework of bifurcation dynamics numerical simulation, the evolution process of specific bifurcation dynamics behavior was gradually visualized to reveal the influence mechanism of the Allee effect and harvesting effort. The research results indicated that the Allee effect and harvesting effort not only seriously affected the bifurcation dynamics essential characteristics of the model (1.3), but also could promote the formation of constant steady state and periodic oscillation persistent survival mode of prey and predator. Furthermore, it is worth noting that appropriate harvesting effort was beneficial for the formation of a sustainable survival cycle between prey and predator. In summary, we hoped that the research findings could contribute to the comprehensive promotion of bifurcation dynamics studies in the predator-prey model.
Citation: Yichao Shao, Hengguo Yu, Chenglei Jin, Jingzhe Fang, Min Zhao. Dynamics analysis of a predator-prey model with Allee effect and harvesting effort[J]. Electronic Research Archive, 2024, 32(10): 5682-5716. doi: 10.3934/era.2024263
In the paper, a predator-prey model with the Allee effect and harvesting effort was proposed to explore the interaction mechanism between prey and predator. Under the framework of mathematical theory deduction, some conditions for the occurrence of transcritical, saddle-node, Hopf, and Bogdanov-Takens bifurcations were derived with harvesting effort and the Allee effect as key parameters. Under the framework of bifurcation dynamics numerical simulation, the evolution process of specific bifurcation dynamics behavior was gradually visualized to reveal the influence mechanism of the Allee effect and harvesting effort. The research results indicated that the Allee effect and harvesting effort not only seriously affected the bifurcation dynamics essential characteristics of the model (1.3), but also could promote the formation of constant steady state and periodic oscillation persistent survival mode of prey and predator. Furthermore, it is worth noting that appropriate harvesting effort was beneficial for the formation of a sustainable survival cycle between prey and predator. In summary, we hoped that the research findings could contribute to the comprehensive promotion of bifurcation dynamics studies in the predator-prey model.

| [1] |
G. B. Gao, D. Bai, T. L. Li, J. Li, Y. Jia, J. Li, et al., Understanding filamentous cyanobacteria and their adaptive niches in Lake Honghu, a shallow eutrophic lake, J. Environ. Sci., 152 (2025), 219–234. https://doi.org/10.1016/j.jes.2024.05.010. doi: 10.1016/j.jes.2024.05.010

|
| [2] |
H. Fang, T. Wu, S. T. Ma, Y. Miao, X. Wang, Biogenic emission as a potential source of atmospheric aromatic hydrocarbons: Insights from a cyanobacterial bloom-occurring eutrophic lake, J. Environ. Sci., 151 (2025), 497–504. https://doi.org/10.1016/j.jes.2024.04.011. doi: 10.1016/j.jes.2024.04.011
|
| [3] |
X. X. Liu, Y. J. Lou, Global dynamics of a predator–prey model, J. Math. Anal. Appl., 371 (2010), 323–340. https://doi.org/10.1016/j.jmaa.2010.05.037. doi: 10.1016/j.jmaa.2010.05.037
|
| [4] |
A. J. Lotka, Elements of physical biology, Nature, 461 (1925). https://doi.org/10.1038/116461b0 doi: 10.1038/116461b0
|
| [5] |
V. Volterra, Fluctuations in the abundance of a species considered mathematically, Nature, 1926 (1926), 558–560. https://doi.org/10.1038/118558a0 doi: 10.1038/118558a0
|
| [6] |
Y. Yao, L. L. Liu, Dynamics of a predator–prey system with foraging facilitation and group defense, Commun. Nonlinear Sci. Numer. Simul., 138 (2024), 108198. https://doi.org/10.1016/j.cnsns.2024.108198. doi: 10.1016/j.cnsns.2024.108198
|
| [7] |
A. A. Thirthar, P. Panja, S. J. Majeed, K. S. Nisar, Dynamic interactions in a two-species model of the mammalian predator–prey system: The influence of Allee effects, prey refuge, water resources, and moonlights, Partial Differ. Equations Appl. Math., 11 (2024), 100865. https://doi.org/10.1016/j.padiff.2024.100865. doi: 10.1016/j.padiff.2024.100865
|
| [8] |
Y. J. Li, M. X. He, Z. Li, Dynamics of a ratio-dependent Leslie–Gower predator–prey model with Allee effect and fear effect, Math. Comput. Simul., 201 (2022), 417–439. https://doi.org/10.1016/j.matcom.2022.05.017. doi: 10.1016/j.matcom.2022.05.017
|
| [9] |
X. Chen, W. Yang, Impact of fear-induced group defense in a Monod–Haldane type prey–predator model, J. Appl. Math. Comput., 212 (2024), 3331–3368. https://doi.org/10.1007/s12190-024-02101-8 doi: 10.1007/s12190-024-02101-8
|
| [10] |
D. Das, T. K. Kar, D. Pal, The impact of invasive species on some ecological services in a harvested predator–prey system, Math. Comput. Simul., 212 (2023), 66–90. https://doi.org/10.1016/j.matcom.2023.04.024. doi: 10.1016/j.matcom.2023.04.024
|
| [11] |
T. K. Ang, H. M. Safuan, Dynamical behaviors and optimal harvesting of an intraguild prey-predator fishery model with Michaelis-Menten type predator harvesting, BioSystems, 202 (2021), 104357. https://doi.org/10.1016/j.biosystems.2021.104357. doi: 10.1016/j.biosystems.2021.104357
|
| [12] |
L. N. Guin, H. Baek, Bistability in modified Holling Ⅱ response model with harvesting and Allee effect: Exploring transitions in a noisy environment, Math. Comput. Simul., 146 (2018), 100–117. https://doi.org/10.1016/j.matcom.2017.10.015. doi: 10.1016/j.matcom.2017.10.015
|
| [13] |
L. N. Guin, S. Djilali, S. Chakravarty, Bistability Cross-diffusion-driven instability in an interacting species model with prey refuge, Chaos, Solitons Fractals, 153 (2021), 111501. https://doi.org/10.1016/j.chaos.2021.111501. doi: 10.1016/j.chaos.2021.111501
|
| [14] |
P. H. Leslie, Some further notes on the use of matrices in population mathematics, Biometrika, 35 (1948), 213–245. https://doi.org/10.2307/2332342 doi: 10.2307/2332342
|
| [15] |
P. H. Leslie, A stochastic model for studying the properties of certain biological systems by numerical methods, Biometrika, 45 (1958), 16–31. https://doi.org/10.1093/biomet/45.1-2.16 doi: 10.1093/biomet/45.1-2.16
|
| [16] |
A. Korobeinikov, A Lyapunov function for Leslie-Gower predator-prey models, Appl. Math. Lett., 14 (2001), 697–699. https://doi.org/10.1016/S0893-9659(01)80029-X. doi: 10.1016/S0893-9659(01)80029-X
|
| [17] |
X. B. Zhang, Q. An, L. Wang, Spatiotemporal dynamics of a delayed diffusive ratio-dependent predator-prey model with fear effect, Nonlinear Dyn., 105 (2021), 3775–3790. https://doi.org/10.1007/s11071-021-06780-x doi: 10.1007/s11071-021-06780-x
|
| [18] |
P. P. Cong, M. Fan, X. F. Zou, Dynamics of a three-species food chain model with fear effect, Commun. Nonlinear Sci. Numer. Simul., 99 (2021), 105809. https://doi.org/10.1016/j.cnsns.2021.105809 doi: 10.1016/j.cnsns.2021.105809
|
| [19] |
Q. Li, Y. Y. Zhang, Y. N. Xiao, Canard phenomena for a slow-fast predator-prey system with group defense of the prey, J. Math. Anal. Appl., 527 (2023), 127418. https://doi.org/10.1016/j.jmaa.2023.127418 doi: 10.1016/j.jmaa.2023.127418
|
| [20] |
D. Mukherjee, C. Maji, Bifurcation analysis of a Holling type Ⅱ predator-prey model with refuge, Chin. J. Phys., 65 (2020), 153–162. https://doi.org/10.1016/j.cjph.2020.02.012 doi: 10.1016/j.cjph.2020.02.012
|
| [21] |
K. Chakraborty, S. S. Das, Biological conservation of a prey-predator system incorporating constant prey refuge through provision of alternative food to predators: a theoretical study, Acta Biotheor., 62 (2014), 183–205. https://doi.org/10.1007/s10441-014-9217-9. doi: 10.1007/s10441-014-9217-9
|
| [22] |
D. Sen, S. Ghorai, S. Sharma, M. Banerjee, Allee effect in prey's growth reduces the dynamical complexity in prey-predator model with generalist predator, Appl. Math. Modell., 91 (2021), 768–790. https://doi.org/10.1016/j.apm.2020.09.046 doi: 10.1016/j.apm.2020.09.046
|
| [23] |
D. Y. Bai, Y. Kang, S. G. Ruan, L. Wang, Dynamics of an intraguild predation food web model with strong Allee effect in the basal prey, Nonlinear Anal. Real World Appl., 58 (2021), 103206. https://doi.org/10.1016/j.nonrwa.2020.103206 doi: 10.1016/j.nonrwa.2020.103206
|
| [24] |
V. Tiwari, J. P. Tripathi, R. K. Upadhyay, Y. P. Wu, J. S. Wang, G. Q. Sun, Predator-prey interaction system with mutually interfering predator: role of feedback control, Appl. Math. Modell., 87 (2022), 222–244. https://doi.org/10.1016/j.apm.2020.04.024 doi: 10.1016/j.apm.2020.04.024
|
| [25] |
P. P. Cong, M. Fan, X. F. Zou, Dynamics of a three-species food chain model with fear effect, Commun. Nonlinear Sci. Numer. Simul., 99 (2021), 105809. https://doi.org/10.1016/j.cnsns.2021.105809 doi: 10.1016/j.cnsns.2021.105809
|
| [26] |
X. X. Li, H. G. Yu, C. J. Dai, Z. Ma, Q. Wang, M. Zhao, Bifurcation analysis of a new aquatic ecological model with aggregation effect, Math. Comput. Simul., 190 (2021), 75–96. https://doi.org/10.1016/j.matcom.2021.05.015 doi: 10.1016/j.matcom.2021.05.015
|
| [27] | W. C. Allee, Animal Aggregations, A Study in General Sociology, The University of Chicago Press, 1431 (1931). https://doi.org/10.5962/bhl.title.7313 |
| [28] |
C. Arancibia–Ibarra, J. Flores, Dynamics of a Leslie–Gower predator–prey model with Holling type Ⅱ functional response, Allee effect and a generalist predator, Math. Comput. Simul., 188 (2021), 1–22. https://doi.org/10.1016/j.matcom.2021.03.035 doi: 10.1016/j.matcom.2021.03.035
|
| [29] |
M. X. He, Z. Li, Global dynamics of a Leslie–Gower predator–prey model with square root response function, Appl. Math. Lett., 140 (2023), 108561. https://doi.org/10.1016/j.aml.2022.108561 doi: 10.1016/j.aml.2022.108561
|
| [30] |
A. Ali, S. Jawad, A. H. Ali, M. Winter, Stability analysis for the phytoplankton-zooplankton model with depletion of dissolved oxygen and strong Allee effects, Results Eng., 22 (2024), 102190. https://doi.org/10.1016/j.rineng.2024.102190 doi: 10.1016/j.rineng.2024.102190
|
| [31] |
S. Akhtar, N. H. Gazi, S. Sarwardi, Mathematical modelling and bifurcation analysis of an eco-epidemiological system with multiple functional responses subjected to Allee effect and competition, Results Control Optim., 15 (2024), 100421. https://doi.org/10.1016/j.rico.2024.100421 doi: 10.1016/j.rico.2024.100421
|
| [32] |
A. Alamin, A. Akgül, M. Rahaman, S. P. Mondal, S. Alam, Dynamical behaviour of discrete logistic equation with Allee effect in an uncertain environment, Results Control Optim., 12 (2023), 100254. https://doi.org/10.1016/j.rico.2023.100254 doi: 10.1016/j.rico.2023.100254
|
| [33] |
S. B. Hsu, T. W. Huang, Global stability for a class of predator-prey systems, SIAM J. Appl. Math, 55 (1995), 763–783. https://doi.org/10.1137/S0036139993253201 doi: 10.1137/S0036139993253201
|
| [34] |
S. J. Lv, Z. M. Fang, The dynamic complexity of a host–parasitoid model with a Beddington-DeAngelis functional response, Chaos, Solitons Fractals, 41 (2009), 2617–2623. https://doi.org/10.1016/j.chaos.2008.09.052 doi: 10.1016/j.chaos.2008.09.052
|
| [35] |
M. Zhao, S. J. Lv, Chaos in a three-species food chain model with a Beddington-DeAngelis functional response, Chaos, Solitons Fractals, 40 (2009), 2305–2316. https://doi.org/10.1016/j.chaos.2007.10.025 doi: 10.1016/j.chaos.2007.10.025
|
| [36] |
S. W. Zhang, L. S. Chen, A study of predator–prey models with the Beddington–DeAnglis functional response and impulsive effect, Chaos, Solitons Fractals, 27 (2006), 237–248. https://doi.org/10.1016/j.chaos.2005.03.039 doi: 10.1016/j.chaos.2005.03.039
|
| [37] |
S. Creel, D. Christianson, Relationships between direct predation and risk effects, Trends Ecol. Evol., 23 (2008), 194–201. https://doi.org/10.1016/j.tree.2007.12.004 doi: 10.1016/j.tree.2007.12.004
|
| [38] |
A. K. Umrao, S. Roy, P. K. Tiwari, P. K. Srivastava, Dynamical behaviors of autonomous and nonautonomous models of generalist predator–prey system with fear, mutual interference and nonlinear harvesting, Chaos, Solitons Fractals, 183 (2024), 114891. https://doi.org/10.1016/j.chaos.2024.114891 doi: 10.1016/j.chaos.2024.114891
|
| [39] |
Y. C. Xu, Y. Yang, F. W. Meng, S. Ruan, Degenerate codimension-2 cusp of limit cycles in a Holling–Tanner model with harvesting and anti-predator behavior, Nonlinear Anal. Real World Appl., 76 (2024), 103995. https://doi.org/10.1016/j.nonrwa.2023.103995 doi: 10.1016/j.nonrwa.2023.103995
|
| [40] |
S. Mandal, N. Sk, P. K. Tiwari, J. Chattopadhyay, Bistability in modified Holling Ⅱ response model with harvesting and Allee effect: Exploring transitions in a noisy environment, Chaos, Solitons Fractals, 178 (2024), 114365. https://doi.org/10.1016/j.chaos.2023.114365 doi: 10.1016/j.chaos.2023.114365
|
| [41] |
X. B. Zhang, H. L. Zhu, Q. An, Dynamics analysis of a diffusive predator-prey model with spatial memory and nonlocal fear effect, J. Math. Anal. Appl., 525 (2023), 127123. https://doi.org/10.1016/j.jmaa.2023.127123 doi: 10.1016/j.jmaa.2023.127123
|
| [42] |
W. J. Li, G. D. Li, J. D. Cao, F. Xu, Dynamics analysis of a diffusive SIRI epidemic system under logistic source and general incidence rate, Commun. Nonlinear Sci. Numer. Simul., 129 (2024), 107675. https://doi.org/10.1016/j.cnsns.2023.107675 doi: 10.1016/j.cnsns.2023.107675
|
| [43] |
J. Z. Cao, H. Y. Sun, P. M. Hao, P. Wang, Bifurcation and turing instability for a predator-prey model with nonlinear reaction cross-diffusion, Appl. Math. Modell., 89 (2021), 1663–-1677. https://doi.org/10.1016/j.apm.2020.08.030 doi: 10.1016/j.apm.2020.08.030
|
| [44] |
X. B. Zhang, Q. An, A. Moussaoui, Effect of density-dependent diffusion on a diffusive predator–prey model in spatially heterogeneous environment, Math. Comput. Simul., 227 (2025), 1–18. https://doi.org/10.1016/j.matcom.2024.07.022 doi: 10.1016/j.matcom.2024.07.022
|
| [45] |
W. J. Li, Y. J. Guan, J. D. Cao, F. Xu, A note on global stability of a degenerate diffusion avian influenza model with seasonality and spatial Heterogeneity, Appl. Math. Lett., 148 (2024), 108884. https://doi.org/10.1016/j.aml.2023.108884 doi: 10.1016/j.aml.2023.108884
|
| [46] |
J. Z. Cao, R. Yuan, Bifurcation analysis in a modified Lesile–Gower model with Holling type Ⅱ functional response and delay, Nonlinear Dyn., 84 (2016), 1341–1352. https://doi.org/10.1007/s11071-015-2572-5 doi: 10.1007/s11071-015-2572-5
|
| [47] |
W. J. Li, Y. Zhang, J. C. Ji, L. Huang, Dynamics of a diffusion epidemic SIRI system in heterogeneous environment, Z. Angew. Math. Phys., 74 (2023), 104. https://doi.org/10.1007/s00033-023-02002-z doi: 10.1007/s00033-023-02002-z
|
| [48] |
W. J. Li, W. R. Zhao, J. D. Cao, L. Huang, Dynamics of a linear source epidemic system with diffusion and media impact, Z. Angew. Math. Phys., 75 (2024), 144. https://doi.org/10.1007/s00033-024-02271-2 doi: 10.1007/s00033-024-02271-2
|
| [49] | L. Perko, Differential Equations and Dynamical Systems, Springer Science and Business Media, 7470 (2013). |




Figures(5)
Yichao Shao, Hengguo Yu, Chenglei Jin, Jingzhe Fang, Min Zhao. Dynamics analysis of a predator-prey model with Allee effect and harvesting effort[J]. Electronic Research Archive, 2024, 32(10): 5682-5716. doi: 10.3934/era.2024263







 DownLoad:
DownLoad: