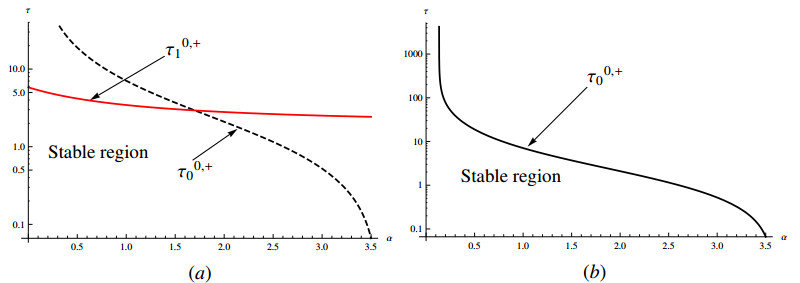
In this paper, a delayed diffusive predator-prey model with the Allee effect and nonlocal competition in prey and hunting cooperation in predators is proposed. The local stability of coexisting equilibrium and the existence of Hopf bifurcation are studied by analyzing the eigenvalue spectrum. The property of Hopf bifurcation is also studied by the center manifold theorem and normal form method. Through numerical simulation, the analysis results are verified, and the influence of these parameters on the model is also obtained. Firstly, increasing the Allee effect parameter $ \beta $ and hunting cooperation parameter $ \alpha $ is not conducive to the stability of the coexistence equilibrium point under some parameters. Secondly, the time delay can also affect the stability of coexisting equilibrium and induce periodic solutions. Thirdly, the nonlocal competition in prey can affect the dynamic properties of the predator-prey model and induce new dynamic phenomena (stably spatially inhomogeneous bifurcating periodic solutions).
Citation: Yujia Xiang, Yuqi Jiao, Xin Wang, Ruizhi Yang. Dynamics of a delayed diffusive predator-prey model with Allee effect and nonlocal competition in prey and hunting cooperation in predator[J]. Electronic Research Archive, 2023, 31(4): 2120-2138. doi: 10.3934/era.2023109
In this paper, a delayed diffusive predator-prey model with the Allee effect and nonlocal competition in prey and hunting cooperation in predators is proposed. The local stability of coexisting equilibrium and the existence of Hopf bifurcation are studied by analyzing the eigenvalue spectrum. The property of Hopf bifurcation is also studied by the center manifold theorem and normal form method. Through numerical simulation, the analysis results are verified, and the influence of these parameters on the model is also obtained. Firstly, increasing the Allee effect parameter $ \beta $ and hunting cooperation parameter $ \alpha $ is not conducive to the stability of the coexistence equilibrium point under some parameters. Secondly, the time delay can also affect the stability of coexisting equilibrium and induce periodic solutions. Thirdly, the nonlocal competition in prey can affect the dynamic properties of the predator-prey model and induce new dynamic phenomena (stably spatially inhomogeneous bifurcating periodic solutions).

| [1] |
Y. Song, Q. Shi, Stability and bifurcation analysis in a diffusive predator-prey model with delay and spatial average, Math. Methods Appl. Sci., 2022. https://doi.org/10.1002/mma.8853 doi: 10.1002/mma.8853

|
| [2] |
C. Xiang, J. Huang, H. Wang, Bifurcations in Holling-Tanner model with generalist predator and prey refuge, J. Differ. Equation, 343 (2023), 495–529. https://doi.org/10.1016/j.jde.2022.10.018 doi: 10.1016/j.jde.2022.10.018
|
| [3] |
K. D. Prasad, B. S. R. V. Prasad, Qualitative analysis of additional food provided predatorprey system with anti-predator behaviour in prey, Nonlinear Dyn., 96 (2019), 1765–1793. https://doi.org/10.1007/s11071-019-04883-0 doi: 10.1007/s11071-019-04883-0
|
| [4] |
J. Zhao, Y. Shao, Bifurcations of a prey-predator system with fear, refuge and additional food, Math. Biosci. Eng., 20 (2023), 3700–3720. http://dx.doi.org/10.3934/mbe.2023173 doi: 10.3934/mbe.2023173
|
| [5] | W. C. Allee, Animal Aggregations, A Study in General Sociology, University of Chicago Press, Chicago, 1931. http://dx.doi.org/10.5962/bhl.title.7313 |
| [6] |
S. Creel, D. Macdonald, Sociality, group size, and reproductive, suppression among carnivores, Advan. Study Behav., 24 (1995), 203–257. http://dx.doi.org/10.1016/S0065-3454(08)60395-2 doi: 10.1016/S0065-3454(08)60395-2
|
| [7] | E. Goodale, G. Beauchamp, G. D. Ruxton, Mixed-Species Groups of Animals: Behavior, Community Structure, and Conservation, Academic Press, 2017. http://dx.doi.org/10.1016/B978-0-12-805355-3.00001-4 |
| [8] |
R. Yadav, N. Mukherjee, M. Sen, Spatiotemporal dynamics of a prey-predator model with Allee effect in prey and hunting cooperation in a Holling type Ⅲ functional response, Nonlinear Dyn., 107 (2022), 1397–1410. https://doi.org/10.1007/s11071-021-07066-y doi: 10.1007/s11071-021-07066-y
|
| [9] |
Y. Song, Y. Peng, T. Zhang, The spatially inhomogeneous Hopf bifurcation induced by memory delay in a memory-based diffusion system, J. Differ. Equation, 300 (2021), 597–624. https://doi.org/10.1016/j.jde.2021.08.010 doi: 10.1016/j.jde.2021.08.010
|
| [10] |
R. Yang, C. Zhang, Dynamics in a diffusive predator-prey system with a constant prey refuge and delay, Nonlinear Anal.-Real World Appl., 31 (2016), 1–22. https://doi.org/10.1016/j.nonrwa.2016.01.005 doi: 10.1016/j.nonrwa.2016.01.005
|
| [11] |
Y. Liu, J. Wei, Double Hopf bifurcation of a diffusive predator-prey system with strong Allee effect and two delays, Nonlinear Anal.-Model Control, 26 (2021), 72–92. https://doi.org/10.15388/namc.2021.26.20561 doi: 10.15388/namc.2021.26.20561
|
| [12] |
R. Yang, D. Jin, W. Wang, A diffusive predator-prey model with generalist predator and time delay. AIMS Math., 7 (2022), 4574–4591. https://doi.org/10.3934/math.2022255 doi: 10.3934/math.2022255
|
| [13] |
R. Yang, X. Zhao, Y. An, Dynamical analysis of a delayed diffusive predator-prey model with additional food provided and anti-predator behavior, Mathematics, 10 (2022), 469. https://doi.org/10.3390/math10030469 doi: 10.3390/math10030469
|
| [14] |
R. Yang, Q. Song, Y. An, Spatiotemporal dynamics in a predator-prey model with functional response increasing in both predator and prey densities, Mathematics, 10 (2022), 17. https://doi.org/10.3390/math10010017 doi: 10.3390/math10010017
|
| [15] |
R. Yang, C. Nie, D. Jin, Spatiotemporal dynamics induced by nonlocal competition in a diffusive predator-prey system with habitat complexity, Nonlinear Dyn., 110 (2022), 879–900. https://doi.org/10.1007/s11071-022-07625-x doi: 10.1007/s11071-022-07625-x
|
| [16] |
R. Yang, F. Wang, D. Jin, Spatially inhomogeneous bifurcating periodic solutions induced by nonlocal competition in a predator-prey system with additional food, Math. Methods Appl. Sci., 45 (2022), 9967–9978. https://doi.org/10.1002/mma.8349 doi: 10.1002/mma.8349
|
| [17] |
N. F. Britton, Aggregation and the competitive exclusion principle, J. Theor. Biol., 136 (1989), 57–66. https://doi.org/10.1016/S0022-5193(89)80189-4 doi: 10.1016/S0022-5193(89)80189-4
|
| [18] |
J. Furter, M. Grinfeld, Local vs. non-local interactions in population dynamics, J. Math. Biol., 27 (1989), 65–80. https://doi.org/10.1007/BF00276081 doi: 10.1007/BF00276081
|
| [19] |
D. Geng, H. Wang, Normal form formulations of double-Hopf bifurcation for partial functional differential equations with nonlocal effect, J. Differ. Equation, 2022 (2022), 741–785. https://doi.org/10.1016/j.jde.2021.11.046 doi: 10.1016/j.jde.2021.11.046
|
| [20] |
D. Geng, W. Jiang, Y. Lou, H. Wang, Spatiotemporal patterns in a diffusive predator-prey system with nonlocal intraspecific prey competition, Stud. Appl. Math., (2021), 1–37. https://doi.org/10.1111/sapm.12444 doi: 10.1111/sapm.12444
|
| [21] |
Y. Liu, D. Duan, B. Niu, Spatiotemporal dynamics in a diffusive predator-prey model with group defense and nonlocal competition, Appl. Math. Lett., 103 (2020), 106175. https://doi.org/10.1016/j.aml.2019.106175 doi: 10.1016/j.aml.2019.106175
|
| [22] | J. Wu, Theory and Applications of Partial Functional Differential Equations, Springer Science Business Media, 1996. https://doi.org/10.1007/978-1-4612-4050-1 |
| [23] | B. D. Hassard, N. D. Kazarinoff, Y. H. Wan, Theory and Applications of Hopf Bifurcation, Cambridge University Press, Cambridge, 1981. http://dx.doi.org/10.1090/conm/445 |






Figures(7)
Yujia Xiang, Yuqi Jiao, Xin Wang, Ruizhi Yang. Dynamics of a delayed diffusive predator-prey model with Allee effect and nonlocal competition in prey and hunting cooperation in predator[J]. Electronic Research Archive, 2023, 31(4): 2120-2138. doi: 10.3934/era.2023109







 DownLoad:
DownLoad: