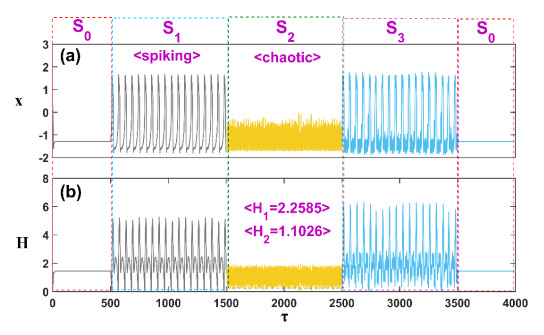
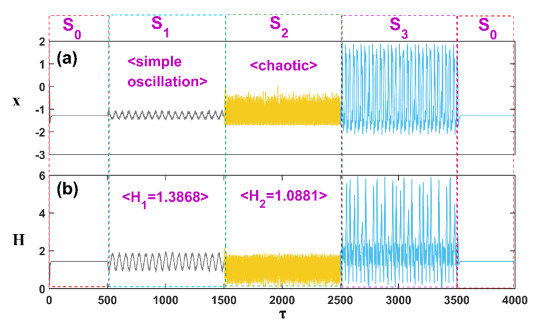
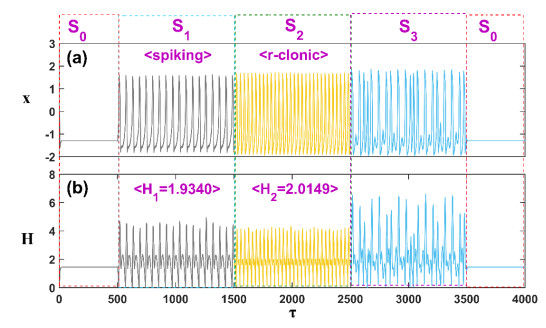
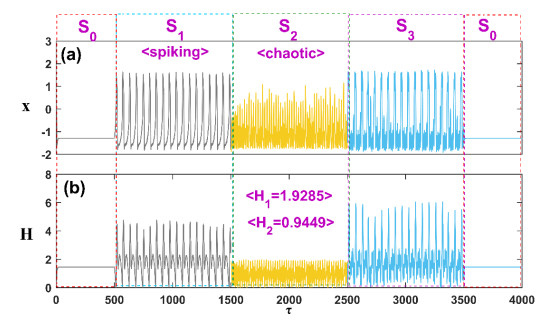
Patients with congenital analgesia who lack the ability to sense temperature generally face low survival rates, highlighting a critical need to understand the underlying mechanisms of heat sensation. While previous research has focused on modeling neural responses to stimuli, the specific mechanisms by which heat-sensitive neurons respond to external temperature changes remain unclear. This gap in knowledge is particularly relevant, as identifying how these neurons react to diverse stimuli can provide insight into sensory deficits linked to congenital analgesia. In this study, we developed a model of heat-sensitive neurons based on the FitzHugh-Nagumo (FHN) neural circuit to investigate neuronal response patterns to external heat stimuli. Two distinct stimulus patterns, each combined with Gaussian white noise, were applied to the model to induce varied firing modes. By calculating the Hamilton energy for each firing mode, we quantified the impact of each external stimulus on neuronal activity. A correlation function was further defined to explore how different stimuli influence the selection of firing modes. Simulation results demonstrate that heat-sensitive neurons show a preferential response to stimuli that induce spike discharge over stimuli that induce r-clonic patterns, as seen in changes to the periodic attractor contours. When exposed to Chua's circuit stimulus, chaotic emission patterns reveal significant shifts in attractor contour, indicating a strong response to spike, r-clonic, and periodic stimuli. These findings suggest that external stimuli capable of inducing spike-and-wave or r-clonic patterns are sensitively detected by thermosensitive neurons, leading to heightened Hamilton energy release and increased regularity in neural activity. This study enhances our understanding of thermosensitive neuronal dynamics under complex stimuli, shedding light on potential response mechanisms relevant to sensory dysfunction in congenital analgesia and advancing the broader field of neural response modeling.
Citation: Yunhai Wang, Guodong Huang, Rui Zhu, Shu Zhou, Yuan Chai. Response mechanism of heat-sensitive neurons under combined noise stimulation[J]. Electronic Research Archive, 2024, 32(11): 6405-6423. doi: 10.3934/era.2024298
Patients with congenital analgesia who lack the ability to sense temperature generally face low survival rates, highlighting a critical need to understand the underlying mechanisms of heat sensation. While previous research has focused on modeling neural responses to stimuli, the specific mechanisms by which heat-sensitive neurons respond to external temperature changes remain unclear. This gap in knowledge is particularly relevant, as identifying how these neurons react to diverse stimuli can provide insight into sensory deficits linked to congenital analgesia. In this study, we developed a model of heat-sensitive neurons based on the FitzHugh-Nagumo (FHN) neural circuit to investigate neuronal response patterns to external heat stimuli. Two distinct stimulus patterns, each combined with Gaussian white noise, were applied to the model to induce varied firing modes. By calculating the Hamilton energy for each firing mode, we quantified the impact of each external stimulus on neuronal activity. A correlation function was further defined to explore how different stimuli influence the selection of firing modes. Simulation results demonstrate that heat-sensitive neurons show a preferential response to stimuli that induce spike discharge over stimuli that induce r-clonic patterns, as seen in changes to the periodic attractor contours. When exposed to Chua's circuit stimulus, chaotic emission patterns reveal significant shifts in attractor contour, indicating a strong response to spike, r-clonic, and periodic stimuli. These findings suggest that external stimuli capable of inducing spike-and-wave or r-clonic patterns are sensitively detected by thermosensitive neurons, leading to heightened Hamilton energy release and increased regularity in neural activity. This study enhances our understanding of thermosensitive neuronal dynamics under complex stimuli, shedding light on potential response mechanisms relevant to sensory dysfunction in congenital analgesia and advancing the broader field of neural response modeling.

| [1] |
S. A. H. Batouli, Seven Ambiguities in explaining the human memory system in the principles of neural science book, Basic Clin. Neurosci., 14 (2023), 543-548. https://doi.org/10.32598/bcn.2023.1774.4 doi: 10.32598/bcn.2023.1774.4

|
| [2] |
S. Masoli, M. F. Rizza, M. Sgritta, W. V. Geit, F. Schürmann, E. D'Angelo, Single neuron optimization as a basis for accurate biophysical modeling: the case of cerebellar granule cells, Front. Cell. Neurosci., 11 (2017), 71. https://doi.org/10.3389/fncel.2017.00071 doi: 10.3389/fncel.2017.00071
|
| [3] |
L. N. Groschner, J. G. Malis, B. Zuidinga, A. Borst, A biophysical account of multiplication by a single neuron, Nature, 603 (2022), 119–123. https://doi.org/10.1038/s41586-022-04428-3 doi: 10.1038/s41586-022-04428-3
|
| [4] |
E. Iavarone, J. Yi, Y. Shi, B. Zandt, C. O'Reilly, W. V. Geit, et al., Experimentally-constrained biophysical models of tonic and burst firing modes in thalamocortical neurons, PLOS Comput. Biol., 15 (2019), e1006753. https://doi.org/10.1371/journal.pcbi.1006753 doi: 10.1371/journal.pcbi.1006753
|
| [5] |
A. R. Cody, B. M. Johnstone, Single auditory neuron response during acute acoustic trauma, Hear. Res., 3 (1980), 3–16. https://doi.org/10.1016/0378-5955(80)90004-0 doi: 10.1016/0378-5955(80)90004-0
|
| [6] |
S. Karak, J. S. Jacobs, M. Kittelmann, C. Spalthoff, R. Katana, E. Sivan-Loukianova, et al., Diverse roles of axonemal dyneins in Drosophila auditory neuron function and mechanical amplification in hearing, Sci. Rep., 5 (2015), 17085. https://doi.org/10.1038/srep17085 doi: 10.1038/srep17085
|
| [7] |
A. Mizrahi, A. Shalev, I. Nelken, Single neuron and population coding of natural sounds in auditory cortex, Curr. Opin. Neurobiol., 24 (2014), 103–110. https://doi.org/10.1016/j.conb.2013.09.007 doi: 10.1016/j.conb.2013.09.007
|
| [8] |
S. Serizawa, K. Miyamichi, H. Sakano, One neuron–one receptor rule in the mouse olfactory system, Trends Genet., 20 (2004), 648–653. https://doi.org/10.1016/j.tig.2004.09.006 doi: 10.1016/j.tig.2004.09.006
|
| [9] |
G. M. Shepherd, Discrimination of molecular signals by the olfactory receptor neuron, Neuron, 13 (1994), 771–790. https://doi.org/10.1016/0896-6273(94)90245-3 doi: 10.1016/0896-6273(94)90245-3
|
| [10] |
A. Menini, Calcium signalling and regulation in olfactory neurons, Curr. Opin. Neurobiol., 9 (1999), 419–426. https://doi.org/10.1016/S0959-4388(99)80063-4 doi: 10.1016/S0959-4388(99)80063-4
|
| [11] |
F. Gabbiani, H. G. Krapp, C. Koch, G. Laurent, Multiplicative computation in a visual neuron sensitive to looming, Nature, 420 (2002), 320–324. https://doi.org/10.1038/nature01190 doi: 10.1038/nature01190
|
| [12] |
S. D. Wiederman, D. C. O'Carroll, Selective attention in an insect visual neuron, Curr. Biol., 23 (2013), 156–161. https://doi.org/10.1016/j.cub.2012.11.048 doi: 10.1016/j.cub.2012.11.048
|
| [13] |
D. A. Butts, Data-driven approaches to understanding visual neuron activity, Ann. Rev. Vis. Sci., 5 (2019), 451–477. https://doi.org/10.1146/annurev-vision-091718-014731 doi: 10.1146/annurev-vision-091718-014731
|
| [14] |
C. Tan, E. K. Cooke, D. E. Leib, Y. Lin, G. E. Daly, C. A. Zimmerman, et al., Warm-sensitive neurons that control body temperature, Cell, 167 (2016), 47–59. https://doi.org/10.1016/j.cell.2016.08.028 doi: 10.1016/j.cell.2016.08.028
|
| [15] | U. Homberg, S. Würden, Movement-sensitive, polarization-sensitive, and light-sensitive neurons of the medulla and accessory medulla of the locust, J. Comp. Neurol., 386 (1997), 329–346. |
| [16] |
E. M. Callaway, R. Yuste, Stimulating neurons with light, Curr. Opin. Neurobiol., 12 (2002), 587–592. https://doi.org/10.1016/S0959-4388(02)00364-1 doi: 10.1016/S0959-4388(02)00364-1
|
| [17] |
M. E. Yamakou, T. D. Tran, L. H. Duc, J. Jost, The stochastic Fitzhugh–Nagumo neuron model in the excitable regime embeds a leaky integrate-and-fire model, J. Math. Biol., 79 (2019), 509–532. https://doi.org/10.1007/s00285-019-01366-z doi: 10.1007/s00285-019-01366-z
|
| [18] |
A. C. Murza, Oscillation patterns in tori of modified FHN neurons, Appl. Math. Modell., 35 (2011), 1096–1106. https://doi.org/10.1016/j.apm.2010.07.055 doi: 10.1016/j.apm.2010.07.055
|
| [19] |
D. Yu, G. Wang, Q. Ding, T. Li, Y. Jia, Effects of bounded noise and time delay on signal transmission in excitable neural networks, Chaos Solitons Fractals, 157 (2022), 111929. https://doi.org/10.1016/j.chaos.2022.111929 doi: 10.1016/j.chaos.2022.111929
|
| [20] |
Y. Jiang, J. Wu, H. Yang, F. Xu, M. Wang, S. Huang, et al., Chimera states mediated by nonlocally attractive-repulsive coupling in FitzHugh–Nagumo neural networks, Chin. J. Phys., 66 (2020), 172–179. https://doi.org/10.1016/j.cjph.2020.03.019 doi: 10.1016/j.cjph.2020.03.019
|
| [21] |
Z. Wu, H. Cheng, Y. Feng, H. Li, Q. Dai, J. Yang, Chimera states in bipartite networks of FitzHugh–Nagumo oscillators, Front. Phys., 13 (2018), 130503. https://doi.org/10.1007/s11467-017-0737-z doi: 10.1007/s11467-017-0737-z
|
| [22] |
F. Yang, Y. Wang, J. Ma, Creation of heterogeneity or defects in a memristive neural network under energy flow, Commun. Nonlinear Sci. Numer. Simul., 119 (2023), 107127. https://doi.org/10.1016/j.cnsns.2023.107127 doi: 10.1016/j.cnsns.2023.107127
|
| [23] |
Y. Wang, C. Wang, G. Ren, J. Tang, W. Jin, Energy dependence on modes of electric activities of neuron driven by multi-channel signals, Nonlinear Dyn., 89 (2017), 1967–1987. https://doi.org/10.1007/s11071-017-3564-4 doi: 10.1007/s11071-017-3564-4
|
| [24] |
K. Usha, P. A. Subha, Collective dynamics and energy aspects of star-coupled Hindmarsh–Rose neuron model with electrical, chemical and field couplings, Nonlinear Dyn., 96(2019), 2115–2124. https://doi.org/10.1007/s11071-019-04909-7 doi: 10.1007/s11071-019-04909-7
|
| [25] |
Y. Yang, J. Ma, Y. Xu, Y. Jia, Energy dependence on discharge mode of Izhikevich neuron driven by external stimulus under electromagnetic induction, Cogn. Neurodyn., 15 (2021), 265–277. https://doi.org/10.1007/s11571-020-09596-4 doi: 10.1007/s11571-020-09596-4
|
| [26] |
B. Ramakrishnan, G. D. Leutcho, K. Rajagopal, S. Jafari, P. L. Ndukum, G. D. Leutcho, Approximate symmetry memristive mega-stable oscillator with attractor growing and its Hamilton energy balance, Eur. Phys. J. Plus, 137 (2022), 596. https://doi.org/10.1140/epjp/s13360-022-02779-4 doi: 10.1140/epjp/s13360-022-02779-4
|
| [27] |
M. Ge, G. Wang, Y. Jia, Influence of the Gaussian colored noise and electromagnetic radiation on the propagation of subthreshold signals in feedforward neural networks, Sci. China Technol. Sci., 64 (2021), 847–857. https://doi.org/10.1007/s11431-020-1696-8 doi: 10.1007/s11431-020-1696-8
|
| [28] |
Y. Chen, L. Yu, Y. Chen, Reliability of weak signals detection in neurons with noise, Sci. China Technol. Sci, 59 (2016), 411–417. https://doi.org/10.1007/s11431-015-6000-3 doi: 10.1007/s11431-015-6000-3
|
| [29] |
P. Fatemeh, R. Karthikeyan, A. Karthikeyan, A. Alsaedi, T. Hayat, V. Pham, Complex dynamics of a neuron model with discontinuous magnetic induction and exposed to external radiation, Cogn. Neurodyn., 12 (2018), 607–614. https://doi.org/10.1007/s11571-018-9497-x doi: 10.1007/s11571-018-9497-x
|
| [30] |
B. Cao, R. Wang, H. Gu, Y. Li, Coherence resonance for neuronal bursting with spike undershoot, Cogn. Neurodyn., 15 (2021), 77–90. https://doi.org/10.1007/s11571-020-09595-5 doi: 10.1007/s11571-020-09595-5
|
| [31] |
Y. Kang, R. Liu, X. Mao, Aperiodic stochastic resonance in neural information processing with Gaussian colored noise, Cogn. Neurodyn., 15 (2021), 517–532. https://doi.org/10.1007/s11571-020-09632-3 doi: 10.1007/s11571-020-09632-3
|
| [32] |
J. Zhao, Y. Qin, Y. Che, H. Ran, J. Li, Effects of network topologies on stochastic resonance in feedforward neural network, Cogn. Neurodyn., 14 (2020), 399–409. https://doi.org/10.1007/s11571-020-09576-8 doi: 10.1007/s11571-020-09576-8
|
| [33] |
Y. Jia, H. Gu, Transition from double coherence resonances to single coherence resonance in a neuronal network with phase noise, Chaos, 25 (2015), 123124. https://doi.org/10.1063/1.4938733 doi: 10.1063/1.4938733
|
| [34] |
X. Yang, Y. Yu, Z. Sun, Autapse-induced multiple stochastic resonances in a modular neuronal network, Chaos, 27 (2017), 083117. https://doi.org/10.1063/1.4999100 doi: 10.1063/1.4999100
|
| [35] |
Y. Xu, M. Liu, Z. Zhu, J. Ma, Dynamics and coherence resonance in a thermosensitive neuron driven by photocurrent, Chin. Phys. B, 29 (2020), 098704. https://doi.org/10.1088/1674-1056/ab9dee doi: 10.1088/1674-1056/ab9dee
|
| [36] |
Y. Yu, F. Han, Q. Wang, Dynamic modeling of neuromodulation techniques: Towards elaboration and individual specificity, Europhysics Letters, 145 (2024), 32001. https://doi.org/10.1209/0295-5075/ad239b doi: 10.1209/0295-5075/ad239b
|
| [37] |
Y. B. Jia, X. L. Yang, J. Kurths, Diversity and time delays induce resonance in a modular neuronal network, Chaos, 24 (2014), 043140. https://doi.org/10.1063/1.4904101 doi: 10.1063/1.4904101
|
| [38] |
Y. Xu, J. Ma, Control of firing activities in thermosensitive neuron by activating excitatory autapse, Chin. Phys. B, 30 (2021), 100501. https://doi.org/10.1088/1674-1056/abeeef doi: 10.1088/1674-1056/abeeef
|
| [39] |
J. Ma, Biophysical neurons, energy, and synapse controllability: A review, J. Zhejiang Univ. Sci. A, 24 (2023), 109–129. https://doi.org/10.1631/jzus.A2200469 doi: 10.1631/jzus.A2200469
|
| [40] |
L. Gammaitoni, P. Hänggi, P. Jung, F. Marchesoni, Stochastic resonance, Rev. Modern Phys., 70 (1998), 223–287. https://doi.org/10.1103/RevModPhys.70.223 doi: 10.1103/RevModPhys.70.223
|
| [41] |
M. Xing, X. Song, Z. Yang, Y. Chen, Bifurcations and excitability in the temperature-sensitive Morris–Lecar neuron, Nonlinear Dyn., 100 (2020), 2687–2698. https://doi.org/10.1007/s11071-020-05667-7 doi: 10.1007/s11071-020-05667-7
|
| [42] |
G. D. Leutcho, L. Woodward, F. Blanchard, Nonlinear dynamics of a single-gap terahertz split-ring resonator under electromagnetic radiation, Chaos, 33 (2023), 103131. https://doi.org/10.1063/5.0157489 doi: 10.1063/5.0157489
|
| [43] |
A. Hariri, M. Babaie-Zadeh, Compressive detection of sparse signals in additive white Gaussian noise without signal reconstruction, Signal Process., 131 (2017), 376–385. https://doi.org/10.1016/j.sigpro.2016.08.020 doi: 10.1016/j.sigpro.2016.08.020
|
| [44] |
Y. Guo, Z. Yao, Y. Xu, J. Ma, Control the stability in chaotic circuit coupled by memristor in different branch circuits, AEU Int. J. Electron. Commun., 145 (2022), 154074. https://doi.org/10.1016/j.aeue.2021.154074 doi: 10.1016/j.aeue.2021.154074
|
| [45] |
Y. Xie, P. Zhou, Z. Yao, J. Ma, Response mechanism in a functional neuron under multiple stimuli, Phys. A, 607 (2022), 128175. https://doi.org/10.1016/j.physa.2022.128175 doi: 10.1016/j.physa.2022.128175
|
| [46] |
N. Kuznetsov, T. Mokaev, V. Ponomarenko, E. Seleznev, N. Stankevich, L. Chua, Hidden attractors in Chua circuit: Mathematical theory meets physical experiments, Nonlinear Dyn., 111 (2023), 5859–5887. https://doi.org/10.1007/s11071-022-08078-y doi: 10.1007/s11071-022-08078-y
|
| [47] |
L. Yan, H. Zhang, Z. Sun, S. Liu, Y. Liu, P. Xiao, Optimization of stimulation waveforms for regulating spike-wave discharges in a thalamocortical model, Chaos Solitons Fractals, 158 (2022), 112025. https://doi.org/10.1016/j.chaos.2022.112025 doi: 10.1016/j.chaos.2022.112025
|
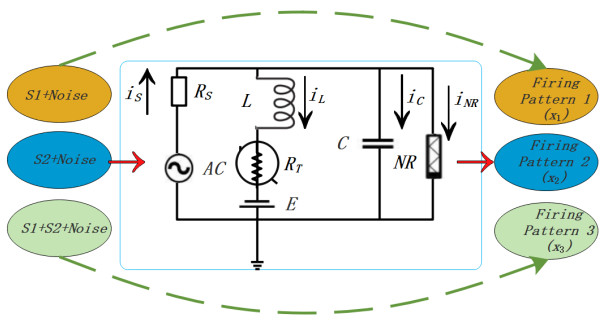
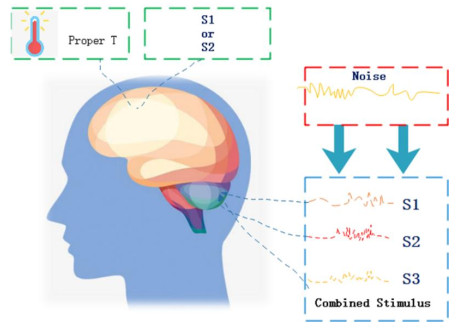
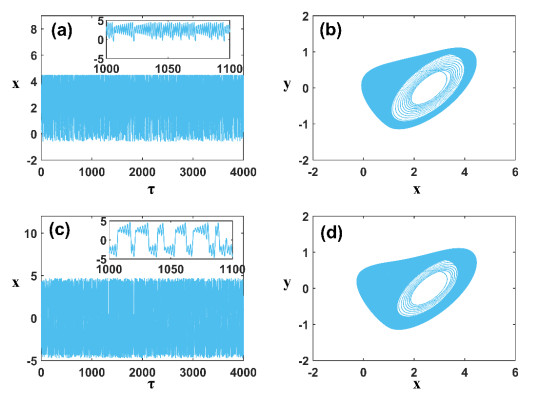
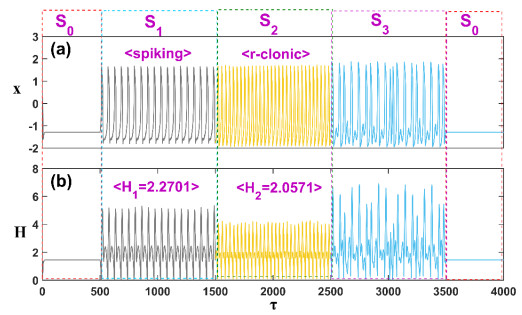
Figures(11)
Yunhai Wang, Guodong Huang, Rui Zhu, Shu Zhou, Yuan Chai. Response mechanism of heat-sensitive neurons under combined noise stimulation[J]. Electronic Research Archive, 2024, 32(11): 6405-6423. doi: 10.3934/era.2024298







 DownLoad:
DownLoad: