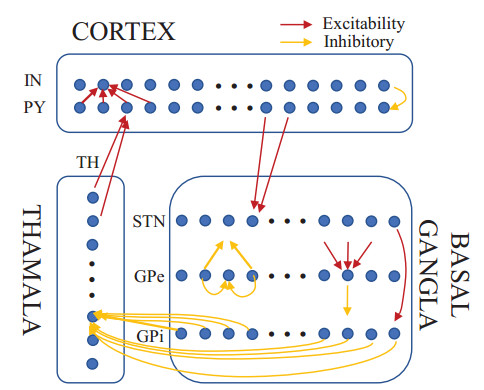
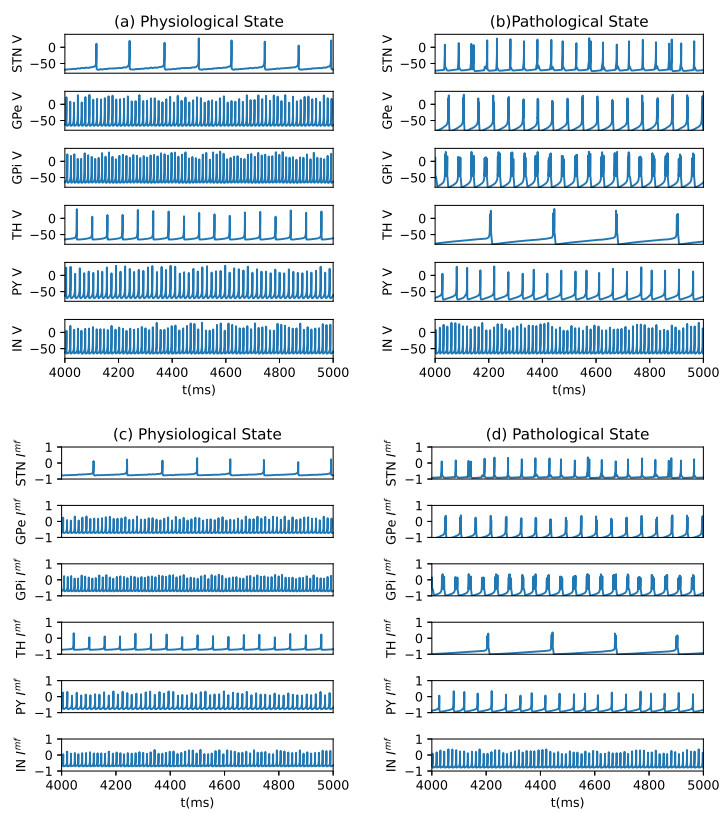
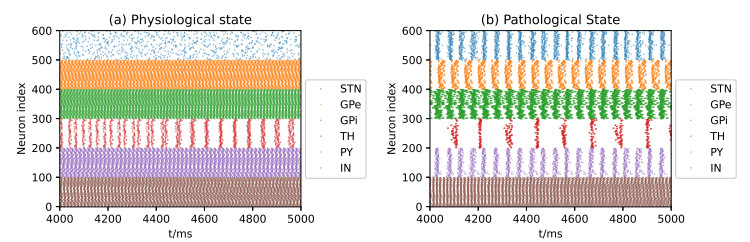
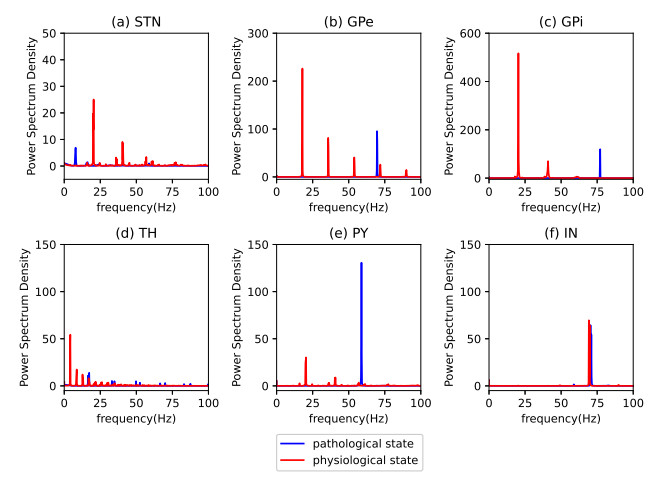
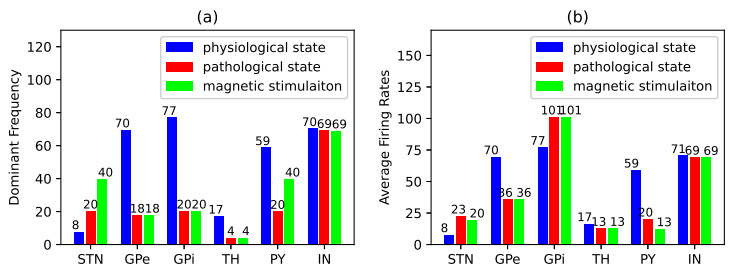
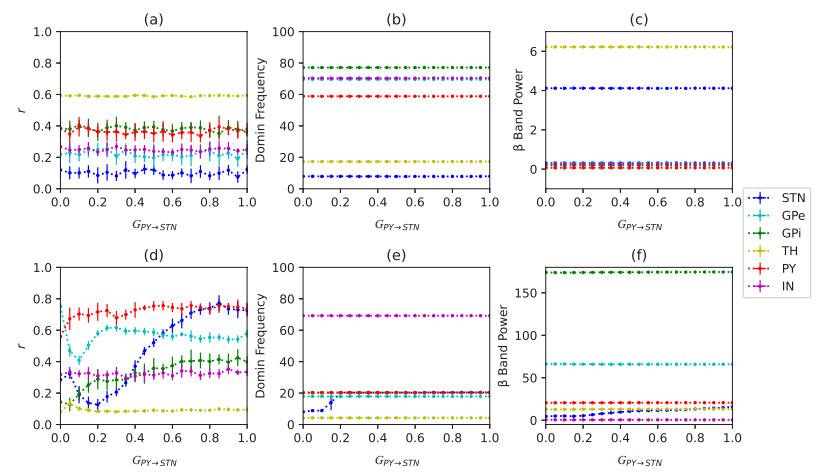
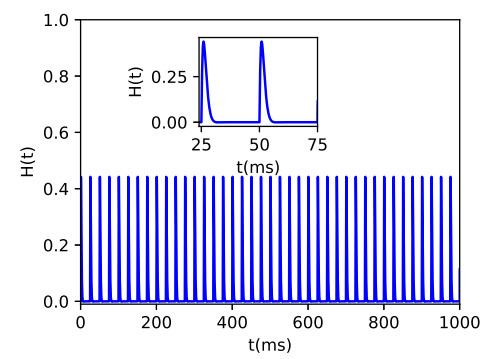
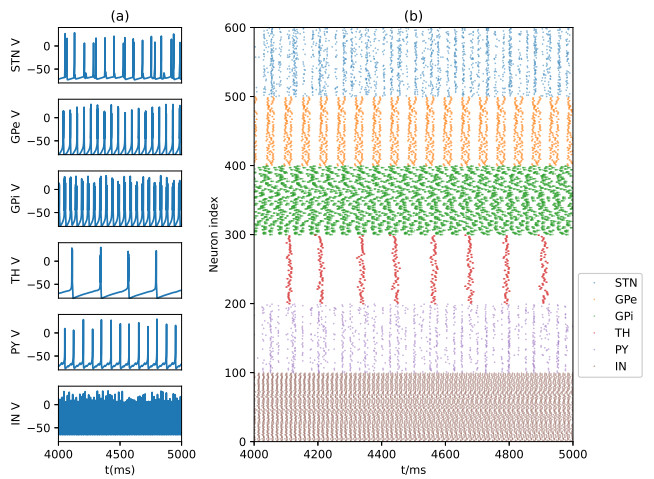
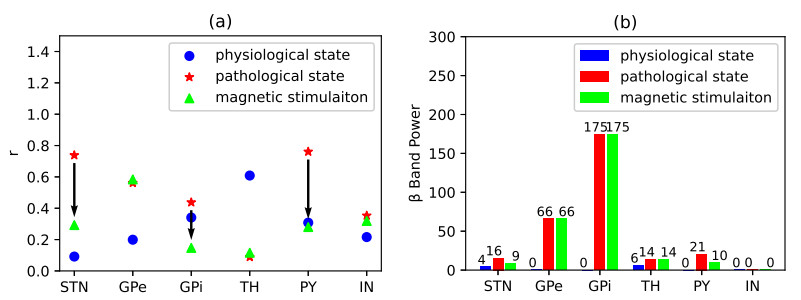
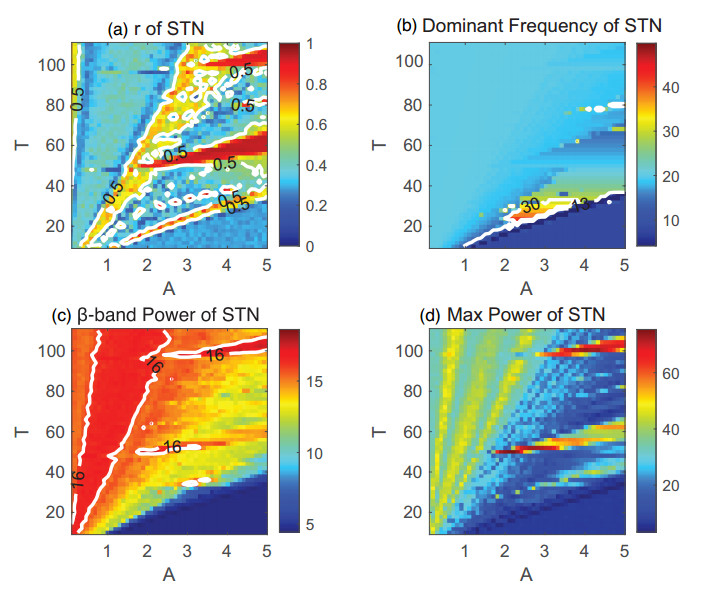
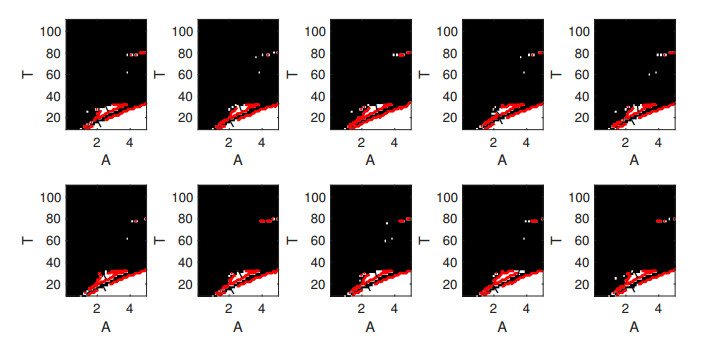
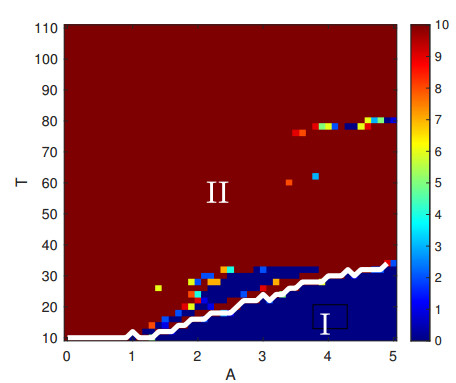
Parkinson's disease (PD) is mainly characterized by changes of firing and pathological oscillations in the basal ganglia (BG). In order to better understand the therapeutic effect of noninvasive magnetic stimulation, which has been used in the treatment of PD, we employ the Izhikevich neuron model as the basic node to study the electrical activity and the controllability of magnetic stimulation in a cortico-basal ganglia-thalamus (CBGT) network. Results show that the firing properties of the physiological and pathological state can be reproduced. Additionally, the electrical activity of pyramidal neurons and strong synapse connection in the hyperdirect pathway cause abnormal $ \beta $-band oscillations and excessive synchrony in the subthalamic nucleus (STN). Furthermore, the pathological firing properties of STN can be efficiently suppressed by external magnetic stimulation. The statistical results give the fitted boundary curves between controllable and uncontrollable regions. This work helps to understand the dynamic response of abnormal oscillation in the PD-related nucleus and provides insights into the mechanisms behind the therapeutic effect of magnetic stimulation.
Citation: Zilu Cao, Lin Du, Honghui Zhang, Lianghui Qu, Luyao Yan, Zichen Deng. Firing activities and magnetic stimulation effects in a Cortico-basal ganglia-thalamus neural network[J]. Electronic Research Archive, 2022, 30(6): 2054-2074. doi: 10.3934/era.2022104
Parkinson's disease (PD) is mainly characterized by changes of firing and pathological oscillations in the basal ganglia (BG). In order to better understand the therapeutic effect of noninvasive magnetic stimulation, which has been used in the treatment of PD, we employ the Izhikevich neuron model as the basic node to study the electrical activity and the controllability of magnetic stimulation in a cortico-basal ganglia-thalamus (CBGT) network. Results show that the firing properties of the physiological and pathological state can be reproduced. Additionally, the electrical activity of pyramidal neurons and strong synapse connection in the hyperdirect pathway cause abnormal $ \beta $-band oscillations and excessive synchrony in the subthalamic nucleus (STN). Furthermore, the pathological firing properties of STN can be efficiently suppressed by external magnetic stimulation. The statistical results give the fitted boundary curves between controllable and uncontrollable regions. This work helps to understand the dynamic response of abnormal oscillation in the PD-related nucleus and provides insights into the mechanisms behind the therapeutic effect of magnetic stimulation.

| [1] |
R. Savica, B. R. Grossardt, J. H. Bower, J. E. Ahlskog, W. A. Rocca, Time trends in the incidence of Parkinson disease, JAMA Neurol., 73 (2016), 981–989. https://doi.org/10.1001/jamaneurol.2016.0947 doi: 10.1001/jamaneurol.2016.0947

|
| [2] |
I. Banegas, I. Prieto, A. Segarra, M. de Gasparo, M. Ramírez-Sánchez, Study of the neuropeptide function in Parkinson's disease using the 6-Hydroxydopamine model of experimental Hemiparkinsonism, AIMS Neurosci., 4 (2017), 223–237. https://doi.org/10.3934/Neuroscience.2017.4.223 doi: 10.3934/Neuroscience.2017.4.223
|
| [3] |
M. G. Krokidis, Identification of biomarkers associated with Parkinson's disease by gene expression profiling studies and bioinformatics analysis, AIMS Neurosci., 6 (2019), 333. https://doi.org/10.3934/Neuroscience.2019.4.333 doi: 10.3934/Neuroscience.2019.4.333
|
| [4] |
P. Vlamos, Novel modeling methodologies for the neuropathological dimensions of Parkinson's disease, AIMS Neurosci., 7 (2020), 89. https://doi.org/10.3934/Neuroscience.2020006 doi: 10.3934/Neuroscience.2020006
|
| [5] |
C. Liu, J. Wang, H. Yu, B. Deng, X. Wei, H. Li, et al., Dynamical analysis of parkinsonian state emulated by hybrid izhikevich neuron models, Commun. Nonlinear Sci. Numer. Simul., 28 (2015), 10–26. https://doi.org/10.1016/j.cnsns.2015.03.018 doi: 10.1016/j.cnsns.2015.03.018
|
| [6] |
H. Bronte-Stewart, C. Barberini, M. M. Koop, B. C. Hill, J. M. Henderson, B. Wingeier, The STN beta-band profile in Parkinson's disease is stationary and shows prolonged attenuation after deep brain stimulation, Exp. Neurol., 215 (2009), 20–28. https://doi.org/10.1016/j.expneurol.2008.09.008 doi: 10.1016/j.expneurol.2008.09.008
|
| [7] |
S. J. van Albada, P. A. Robinson, Mean-field modeling of the basal ganglia-thalamocortical system. I: Firing rates in healthy and parkinsonian states, J. Theor. Biol., 257 (2009), 642–663. https://doi.org/10.1016/j.jtbi.2008.12.018 doi: 10.1016/j.jtbi.2008.12.018
|
| [8] |
Y. Yu, X. Wang, Q. Wang, Q. Wang, A review of computational modeling and deep brain stimulation: applications to Parkinson's disease, Appl. Math. Mech., 41 (2020), 1747–1768. https://doi.org/10.1007/s10483-020-2689-9 doi: 10.1007/s10483-020-2689-9
|
| [9] |
H. Zhang, Y. Yu, Z. Deng, Q. Wang, Activity pattern analysis of the subthalamopallidal network under Channelrhodopsin-2 and Halorhodopsin photocurrent control, Chaos Soliton Fract., 138 (2020), 109963. https://doi.org/10.1016/j.chaos.2020.109963 doi: 10.1016/j.chaos.2020.109963
|
| [10] |
L. Doyle Gaynor, A. Kühn, M. Dileone, V. Litvak, A. Eusebio, A. Pogosyan, et al., Suppression of beta oscillations in the subthalamic nucleus following cortical stimulation in humans, Eur. J. Neurol., 28 (2008), 1686–1695. https://doi.org/10.1111/j.1460-9568.2008.06363.x doi: 10.1111/j.1460-9568.2008.06363.x
|
| [11] |
D. J. Ellens, D. K. Leventhal, electrophysiology of basal ganglia and cortex in models of Parkinson disease, J. Parkinson's Disease, 3 (2013), 241–254. https://doi.org/10.3233/JPD-130204 doi: 10.3233/JPD-130204
|
| [12] |
A. Leblois, T. Boraud, W. Meissner, H. Bergman, D. Hansel, Competition between feedback loops underlies normal and pathological dynamics in the basal ganglia, J. Neurosci., 26 (2006), 3567–3583. https://doi.org/10.1523/JNEUROSCI.5050-05.2006 doi: 10.1523/JNEUROSCI.5050-05.2006
|
| [13] |
A. Pavlides, S. J. Hogan, R. Bogacz, Computational models describing possible mechanisms for generation of excessive beta oscillations in Parkinson's disease, PLoS Comput. Biol., 11 (2015), e1004609. https://doi.org/10.1371/journal.pcbi.1004609 doi: 10.1371/journal.pcbi.1004609
|
| [14] |
M. Lu, X. Wei, K. A. Loparo, Investigating synchronous oscillation and deep brain stimulation treatment in a model of cortico-basal ganglia network, IEEE Trans. Neural Syst. Rehabilitation Eng., 25 (2017), 1950–1958. https://doi.org/10.1109/TNSRE.2017.2707100 doi: 10.1109/TNSRE.2017.2707100
|
| [15] |
P. Davila-Pérez, A. Pascual-Leone, J. Cudeiro, Effects of transcranial static magnetic stimulation on motor cortex evaluated by different TMS waveforms and current directions, Neuroscience, 413 (2019), 22–30. https://doi.org/10.1016/j.neuroscience.2019.05.065 doi: 10.1016/j.neuroscience.2019.05.065
|
| [16] |
M. Lv, J. Ma, Multiple modes of electrical activities in a new neuron model under electromagnetic radiation, Neurocomputing, 205 (2016), 375–381. https://doi.org/10.1016/j.neucom.2016.05.004 doi: 10.1016/j.neucom.2016.05.004
|
| [17] |
C. Yang, Z. Liu, Q. Wang, G. Luan, F. Zhai, Epileptic seizures in a heterogeneous excitatory network with short-term plasticity, Cogn. Neurodyn., 15 (2021), 43–51. https://doi.org/10.1007/s11571-020-09582-w doi: 10.1007/s11571-020-09582-w
|
| [18] |
J. Zhao, D. Fan, Q. Wang, Q. Wang, Dynamical transitions of the coupled class I (II) neurons regulated by an astrocyte, Nonlinear Dyn., 103 (2021), 913–924. https://doi.org/10.1007/s11071-020-06122-3 doi: 10.1007/s11071-020-06122-3
|
| [19] |
M. Stimberg, R. Brette, D. F. Goodman, Brian 2, an intuitive and efficient neural simulator, Elife, 8 (2019), e47314. https://doi.org/10.7554/eLife.47314 doi: 10.7554/eLife.47314
|
| [20] |
C. V. Rusu, M. Murakami, U. Ziemann, J. Triesch, A model of TMS-induced I-waves in motor cortex, Brain Stimul., 7 (2014), 401–414. https://doi.org/10.1016/j.brs.2014.02.009 doi: 10.1016/j.brs.2014.02.009
|
Figures(12) / Tables(4)
Zilu Cao, Lin Du, Honghui Zhang, Lianghui Qu, Luyao Yan, Zichen Deng. Firing activities and magnetic stimulation effects in a Cortico-basal ganglia-thalamus neural network[J]. Electronic Research Archive, 2022, 30(6): 2054-2074. doi: 10.3934/era.2022104







 DownLoad:
DownLoad: