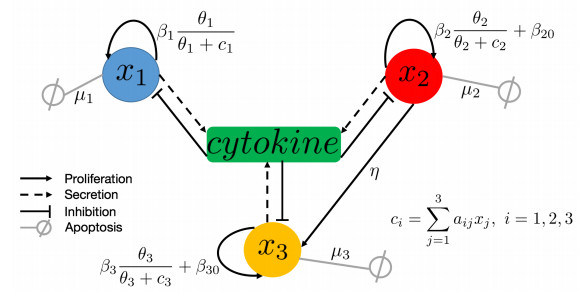
Intratumor heterogeneity hinders the success of anti-cancer treatment due to the interaction between different types of cells. To recapitulate the communication of different types of cells, we developed a mathematical model to study the dynamic interaction between normal, drug-sensitive and drug-resistant cells in response to cancer treatment. Based on the proposed model, we first study the analytical conclusions, namely the nonnegativity and boundedness of solutions, and the existence and stability of steady states. Furthermore, to investigate the optimal treatment that minimizes both the cancer cells count and the total dose of drugs, we apply the Pontryagin's maximum(or minimum) principle (PMP) to explore the combination therapy strategy with either quadratic control or linear control functionals. We establish the existence and uniqueness of the quadratic control problem, and apply the forward-backward sweep method (FBSM) to solve the optimal control problems and obtain the optimal therapy scheme.
Citation: Haifeng Zhang, Jinzhi Lei. Optimal treatment strategy of cancers with intratumor heterogeneity[J]. Mathematical Biosciences and Engineering, 2022, 19(12): 13337-13373. doi: 10.3934/mbe.2022625
Intratumor heterogeneity hinders the success of anti-cancer treatment due to the interaction between different types of cells. To recapitulate the communication of different types of cells, we developed a mathematical model to study the dynamic interaction between normal, drug-sensitive and drug-resistant cells in response to cancer treatment. Based on the proposed model, we first study the analytical conclusions, namely the nonnegativity and boundedness of solutions, and the existence and stability of steady states. Furthermore, to investigate the optimal treatment that minimizes both the cancer cells count and the total dose of drugs, we apply the Pontryagin's maximum(or minimum) principle (PMP) to explore the combination therapy strategy with either quadratic control or linear control functionals. We establish the existence and uniqueness of the quadratic control problem, and apply the forward-backward sweep method (FBSM) to solve the optimal control problems and obtain the optimal therapy scheme.

| [1] |
A. Marusyk, M. Janiszewska, K. Polyak, Intratumor heterogeneity: the rosetta stone of therapy resistance, Cancer Cell, 37 (2020), 471–484. https://doi.org/10.1016/j.ccell.2020.03.007 doi: 10.1016/j.ccell.2020.03.007

|
| [2] |
I. Dagogo-Jack, A. T. Shaw, Tumour heterogeneity and resistance to cancer therapies, Nat. Rev. Clin. Oncol., 15 (2018), 81–94. https://doi.org/10.1038/nrclinonc.2017.166 doi: 10.1038/nrclinonc.2017.166
|
| [3] |
M. Labrie, J. S. Brugge, G. B. Mills, I. K. Zervantonakis, Therapy resistance: opportunities created by adaptive responses to targeted therapies in cancer, Nat. Rev. Cancer, 22 (2022), 323–339. https://doi.org/10.1038/s41568-022-00454-5 doi: 10.1038/s41568-022-00454-5
|
| [4] |
A. N. Hata, M. J. Niederst, H. L. Archibald, M. Gomez-Caraballo, F. M. Siddiqui, H. E. Mulvey, et al., Tumor cells can follow distinct evolutionary paths to become resistant to epidermal growth factor receptor inhibition, Nat. Med., 22 (2016), 262–269. https://doi.org/10.1038/nm.4040 doi: 10.1038/nm.4040
|
| [5] |
C. Roche-Lestienne, V. Soenen-Cornu, N. Grardel-Duflos, J. L. Laï, N. Philippe, T. Facon, et al., Several types of mutations of the Abl gene can be found in chronic myeloid leukemia patients resistant to STI571, and they can pre-exist to the onset of treatment, Blood, 100 (2002), 1014–1018. https://doi.org/10.1182/blood.V100.3.1014 doi: 10.1182/blood.V100.3.1014
|
| [6] |
T. N. Wong, G. Ramsingh, A. L. Young, C. A. Miller, W. Touma, J. S. Welch, et al., Role of TP53 mutations in the origin and evolution of therapy-related acute myeloid leukaemia, Nature, 518 (2015), 552–555. https://doi.org/10.1038/nature13968 doi: 10.1038/nature13968
|
| [7] |
S. Misale, R. Yaeger, S. Hobor, E. Scala, M. Janakiraman, D. Liska, et al., Emergence of KRAS mutations and acquired resistance to anti-EGFR therapy in colorectal cancer, Nature, 486 (2012), 532–536. https://doi.org/10.1038/nature11156 doi: 10.1038/nature11156
|
| [8] |
A. B. Turke, K. Zejnullahu, Y. L. Wu, Y. Song, D. Dias-Santagata, E. Lifshits, et al., Preexistence and clonal selection of MET amplification in EGFR mutant NSCLC, Cancer Cell, 17 (2010), 77–88. https://doi.org/10.1016/j.ccr.2009.11.022 doi: 10.1016/j.ccr.2009.11.022
|
| [9] |
L. A. Diaz, R. T. Williams, J. Wu, I. Kinde, J. R. Hecht, J. Berlin, et al., The molecular evolution of acquired resistance to targeted EGFR blockade in colorectal cancers, Nature, 486 (2012), 537–540. https://doi.org/10.1038/nature11219 doi: 10.1038/nature11219
|
| [10] |
S. Maheswaran, L. V. Sequist, S. Nagrath, L. Ulkus, B. Brannigan, C. V. Collura, et al., Detection of mutations in EGFR in circulating lung-cancer cells, N. Engl. J. Med., 359 (2008), 366–377. https://doi.org/10.1056/NEJMoa0800668 doi: 10.1056/NEJMoa0800668
|
| [11] |
W. Pao, V. A. Miller, K. A. Politi, G. J. Riely, R. Somwar, M. F. Zakowski, et al., Acquired resistance of lung adenocarcinomas to gefitinib or erlotinib is associated with a second mutation in the EGFR kinase domain, PLos Med., 2 (2005), 225–235. https://doi.org/10.1371/journal.pmed.0020073 doi: 10.1371/journal.pmed.0020073
|
| [12] |
M. Russo, G. Crisafulli, A. Sogari, N. M. Reilly, S. Arena, S. Lamba, et al., Adaptive mutability of colorectal cancers in response to targeted therapies, Science, 366 (2019), 1473–1480. https://doi.org/10.1126/science.aav4474 doi: 10.1126/science.aav4474
|
| [13] |
L. Ding, T. J. Ley, D. E. Larson, C. A. Miller, D. C. Koboldt, J. S. Welch, et al., Clonal evolution in relapsed acute myeloid leukaemia revealed by whole-genome sequencing, Nature, 481 (2012), 506–510. https://doi.org/10.1038/nature10738 doi: 10.1038/nature10738
|
| [14] |
Y. Alwash, J. D. Khoury, M. Tashakori, R. Kanagal-Shamanna, N. Daver, F. Ravandi, et al., Development of TP53 mutations over the course of therapy for acute myeloid leukemia, Am. J. Hematol., 96 (2021), 1420–1428. https://doi.org/10.1002/ajh.26314 doi: 10.1002/ajh.26314
|
| [15] |
J. A. Engelman, K. Zejnullahu, T. Mitsudomi, Y. C. Song, C. Hyland, J. O. Park, et al., MET amplification leads to gefitinib resistance in lung cancer by activating ERBB3 signaling, Science, 316 (2007), 1039–1043. https://doi.org/10.1126/science.1141478 doi: 10.1126/science.1141478
|
| [16] |
M. E. Gorre, M. Mohammed, K. Ellwood, N. Hsu, R. Paquette, P. N. Rao, et al., Clinical resistance to STI-571 cancer therapy caused by BCR-ABL gene mutation or amplification, Science, 293 (2001), 876–880. https://doi.org/10.1126/science.1062538 doi: 10.1126/science.1062538
|
| [17] |
J. J. Cunningham, J. S. Brown, R. A. Gatenby, K. Staňková, Optimal control to develop therapeutic strategies for metastatic castrate resistant prostate cancer, J. Theor. Biol., 459 (2018), 67–78. https://doi.org/10.1016/j.jtbi.2018.09.022 doi: 10.1016/j.jtbi.2018.09.022
|
| [18] |
E. Kozłowska, R. Suwiński, M. Giglok, A. Świerniak, M. Kimmel, Mathematical model predicts response to chemotherapy in advanced non-resectable non-small cell lung cancer patients treated with platinum-based doublet, PLoS Comput. Biol., 16 (2020), e1008234. https://doi.org/10.1371/journal.pcbi.1008234 doi: 10.1371/journal.pcbi.1008234
|
| [19] |
A. Y. Yin, J. G. C. van Hasselt, H. J. Guchelaar, L. E. Friberg, D. J. A. R. Moes, Anti-cancer treatment schedule optimization based on tumor dynamics modelling incorporating evolving resistance, Sci Rep, 12 (2022), 4206. https://doi.org/10.1038/s41598-022-08012-7 doi: 10.1038/s41598-022-08012-7
|
| [20] |
J. W. Zhou, Y. T. Liu, Y. B. Zhang, Q. F. Li, Y. G. Cao, Modeling tumor evolutionary dynamics to predict clinical outcomes for patients with metastatic colorectal cancer: a retrospective analysis, Cancer Res., 80 (2020), 591–601. https://doi.org/10.1158/0008-5472.CAN-19-1940 doi: 10.1158/0008-5472.CAN-19-1940
|
| [21] |
N. Picco, E. Sahai, P. K. Maini, A. R. A. Anderson, Integrating models to quantify environment-mediated drug resistance, Cancer Res., 77 (2017), 5409–5418. https://doi.org/10.1158/0008-5472.CAN-17-0835 doi: 10.1158/0008-5472.CAN-17-0835
|
| [22] |
T. Hähnel, C. Baldow, J. Guilhot, F. Guilhot, S. Saussele, S. Mustjoki, et al., Model-based inference and classification of immunologic control mechanisms from TKI cessation and dose reduction in patients with CML, Cancer Res., 80 (2020), 2394–2406. https://doi.org/10.1158/0008-5472.CAN-19-2175 doi: 10.1158/0008-5472.CAN-19-2175
|
| [23] |
E. Piretto, M. Delitala, M. Ferraro, Combination therapies and intra-tumoral competition: insights from mathematical modeling, J. Theor. Biol., 446 (2018), 149–159. https://doi.org/10.1016/j.jtbi.2018.03.014 doi: 10.1016/j.jtbi.2018.03.014
|
| [24] |
S. Sameen, R. Barbuti, P. Milazzo, A. Cerone, M. Del Re, R. Danesi, Mathematical modeling of drug resistance due to KRAS mutation in colorectal cancer, J. Theor. Biol., 389 (2016), 263–273. https://doi.org/10.1016/j.jtbi.2015.10.019 doi: 10.1016/j.jtbi.2015.10.019
|
| [25] |
S. Brown, C. M. Pineda, T. C. Xin, J. Boucher, K. C. Suozzi, et al., Correction of aberrant growth preserves tissue homeostasis, Nature, 548 (2017), 334–337. https://doi.org/10.1038/nature23304 doi: 10.1038/nature23304
|
| [26] |
M. Vishwakarma, E. Piddini, Outcompeting cancer, Nat. Rev. Cancer, 20 (2020), 187–198. https://doi.org/10.1038/s41568-019-0231-8 doi: 10.1038/s41568-019-0231-8
|
| [27] | G. W. Swan, T. L. Vincent, Optimal control analysis in the chemotherapy of IgG multiple myeloma, Bull. Math. Biol., 39 (1977), 317–337. |
| [28] |
F. Castiglione, B. Piccoll, Cancer immunotherapy, mathematical modeling and optimal control, J. Theor. Biol., 247 (2007), 723–732. https://doi.org/10.1016/j.jtbi.2007.04.003 doi: 10.1016/j.jtbi.2007.04.003
|
| [29] |
T. Burden, J. Ernstberger, K. R. Fister, Optimal control applied to immunotherapy, Discrete Contin. Dyn. Syst.-Ser. B, 4 (2004), 135–146. https://doi.org/10.3934/dcdsb.2004.4.135 doi: 10.3934/dcdsb.2004.4.135
|
| [30] |
K. R. Fister, J. C. Panetta, Optimal control applied to competing chemotherapeutic cell-kill strategies, SIAM J. Appl. Math., 63 (2003), 1954–1971. https://doi.org/10.1137/S0036139902413489 doi: 10.1137/S0036139902413489
|
| [31] |
J. A. Sharp, A. P. Browning, T. Mapder, C. M. Baker, K. Burrage, M. J. Simpson, Designing combination therapies using multiple optimal controls, J. Theor. Biol., 497 (2020), 110277. https://doi.org/10.1016/j.jtbi.2020.110277 doi: 10.1016/j.jtbi.2020.110277
|
| [32] |
H. Moore, L. Strauss, U. Ledzewicz, Optimization of combination therapy for chronic myeloid leukemia with dosing constraints, J. Math. Biol., 77 (2018), 1533–1561. https://doi.org/10.1007/s00285-018-1262-6 doi: 10.1007/s00285-018-1262-6
|
| [33] |
U. Ledzewicz, H. Schättler, Combination of antiangiogenic treatment with chemotherapy as a multi-input optimal control problem, Math. Meth. Appl. Sci., 45 (2021), 3058–3082. https://doi.org/10.1002/mma.7977 doi: 10.1002/mma.7977
|
| [34] |
A. Camacho, S. Jerez, Bone metastasis treatment modeling via optimal control, J. Math. Biol., 78 (2019), 497–526. https://doi.org/10.1007/s00285-018-1281-3 doi: 10.1007/s00285-018-1281-3
|
| [35] | L. S. Pontryagin, V. G. Boltyanskii, R. V. Gamkrelidze, E. F. Mishchenko, The mathematical theory of optimal processes, J. Oper. Res. Soc., 16 (1965), 493–494. |
| [36] |
J. Massagué, TGF$\beta$ signalling in context, Nat. Rev. Mol. Cell Biol., 13 (2012), 616–630. https://doi.org/10.1038/nrm3434 doi: 10.1038/nrm3434
|
| [37] |
A. Nakao, M. Afrakhte, A. Morén, T. Nakayama, J. L. Christian, R. Heuchel, et al., Identification of Smad7, a TGF $\beta$-inducible antagonist of TGF-$\beta$ signalling, Nature, 389 (1997), 631–635. https://doi.org/10.1038/39369 doi: 10.1038/39369
|
| [38] |
D. M. Ornitz, N. Itoh, Fibroblast growth factors, Genome Biol., 2 (2001), reviews3005.1–3005.12. https://doi.org/10.1186/gb-2001-2-3-reviews3005 doi: 10.1186/gb-2001-2-3-reviews3005
|
| [39] |
J. Lei, A general mathematical framework for understanding the behavior of heterogeneous stem cell regeneration, J. Theor. Biol., 492 (2020), 110196. https://doi.org/10.1016/j.jtbi.2020.110196 doi: 10.1016/j.jtbi.2020.110196
|
| [40] |
S. Bernard, J. Bélair, M. C. Mackey, Oscillations in cyclical neutrophenia: new evidence based on mathematical modeling, J. Theor. Biol., 223 (2003), 283–298. https://doi.org/10.1016/S0022-5193(03)00090-0 doi: 10.1016/S0022-5193(03)00090-0
|
| [41] |
D. Hanahan, R. A. Weinberg, The hallmarks of cancer, Cell, 100 (2000), 57–70. https://doi.org/10.1016/S0092-8674(00)81683-9 doi: 10.1016/S0092-8674(00)81683-9
|
| [42] | J. K. Hale, Ordinary Differential Equations, Robert E. Krieger Publishing Company, Inc, New York, 1980. |
| [43] | W. H. Fleming, R. W. Rishel, Deterministic and Stochastic Optimal Control, Springer-Verlag, New York, 1975. |
| [44] | A. Bressan, B. Piccoli, Introduction to the Mathematical Theory of Control, American Institute of Mathematical Sciences, 2007. |
| [45] | S. Lenhart, J. T. Workman, Optimal Control Applied to Biological Models, Chapman & Hall/CRC, Taylor & Francis, London, 2007. https://doi.org/10.1201/9781420011418 |






Figures(7) / Tables(1)

Haifeng Zhang, Jinzhi Lei. Optimal treatment strategy of cancers with intratumor heterogeneity[J]. Mathematical Biosciences and Engineering, 2022, 19(12): 13337-13373. doi: 10.3934/mbe.2022625







 DownLoad:
DownLoad: