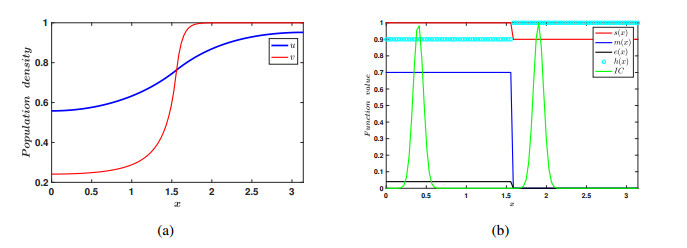
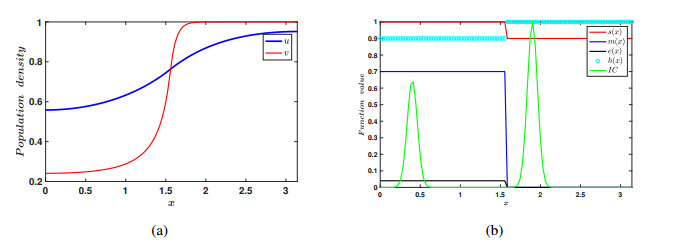
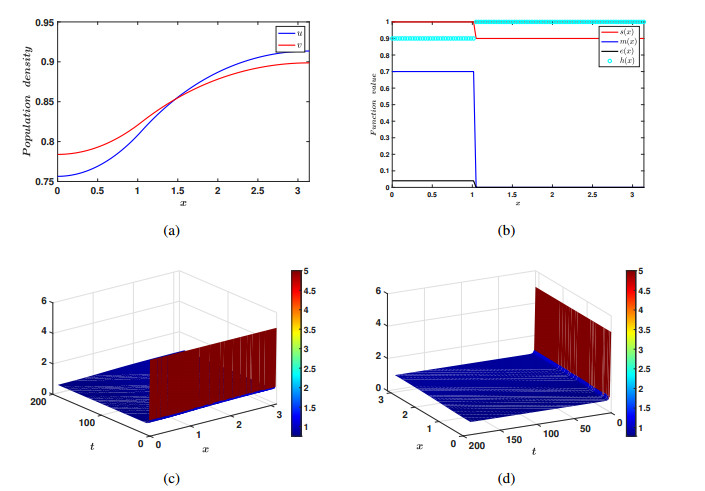
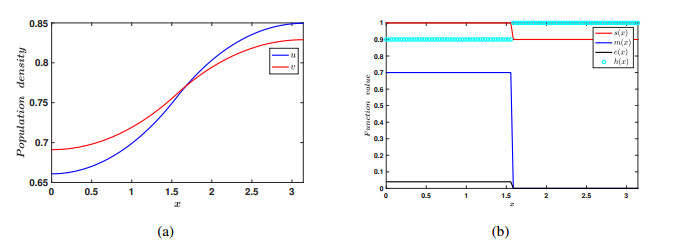
In the current manuscript, a two-patch model with the Allee effect and nonlinear dispersal is presented. We study both the ordinary differential equation (ODE) case and the partial differential equation (PDE) case here. In the ODE model, the stability of the equilibrium points and the existence of saddle-node bifurcation are discussed. The phase diagram and bifurcation curve of our model are also given as a results of numerical simulation. Besides, the corresponding linear dispersal case is also presented. We show that, when the Allee effect is large, high intensity of linear dispersal is not favorable to the persistence of the species. We further show when the Allee effect is large, nonlinear diffusion is more beneficial to the survival of the population than linear diffusion. Moreover, the results of the PDE model extend our findings from discrete patches to continuous patches.
Citation: Yue Xia, Lijuan Chen, Vaibhava Srivastava, Rana D. Parshad. Stability and bifurcation analysis of a two-patch model with the Allee effect and dispersal[J]. Mathematical Biosciences and Engineering, 2023, 20(11): 19781-19807. doi: 10.3934/mbe.2023876
In the current manuscript, a two-patch model with the Allee effect and nonlinear dispersal is presented. We study both the ordinary differential equation (ODE) case and the partial differential equation (PDE) case here. In the ODE model, the stability of the equilibrium points and the existence of saddle-node bifurcation are discussed. The phase diagram and bifurcation curve of our model are also given as a results of numerical simulation. Besides, the corresponding linear dispersal case is also presented. We show that, when the Allee effect is large, high intensity of linear dispersal is not favorable to the persistence of the species. We further show when the Allee effect is large, nonlinear diffusion is more beneficial to the survival of the population than linear diffusion. Moreover, the results of the PDE model extend our findings from discrete patches to continuous patches.

| [1] | M. Luo, S. Wang, S. Saavedra, D. Ebert, F. Altermatt, Multispecies coexistence in fragmented landscapes, in Proceedings of the National Academy of Sciences, (2022), e2201503119. https://doi.org/10.1073/pnas.2201503119 |
| [2] |
M. E. Soule, D. Simberloff, What do genetics and ecology tell us about the design of nature reserves?, Biol. Conserv., 35 (1986), 19–40. https://doi.org/10.1016/0006-3207(86)90025-X doi: 10.1016/0006-3207(86)90025-X

|
| [3] |
R. Channell, M. Lomolino, Dynamic biogeography and conservation of endangered species, Nature, 403 (2000), 84–86. https://doi.org/10.1038/47487 doi: 10.1038/47487
|
| [4] | A. B. Franklin, B. R. Noon, T. L. George, What is habitat fragmentation?, Stud. Avian Biol., 25 (2002), 20–29. |
| [5] | N. Keyghobadi, The genetic implications of habitat fragmentation for animals, Can. J. Zool., 85 (2007), 1049–1064. |
| [6] | L. Fahrig, Effect of habitat fragmentation on the extinction threshold: A synthesis, Ecol. Appl., 12 (2002), 346–353. |
| [7] |
L. Fahrig, Ecological responses to habitat fragmentation per se, Ann. Rev. Ecol. Evol. Syst., 48 (2017), 1–23. https://doi.org/10.1146/annurev-ecolsys-110316-022612 doi: 10.1146/annurev-ecolsys-110316-022612
|
| [8] |
L. Fahrig, Habitat fragmentation: A long and tangled tale, Global Ecol. Biogeography, 28 (2019), 33–41. https://doi.org/10.1111/geb.12839 doi: 10.1111/geb.12839
|
| [9] | M. S. Rohwäder, F. Jeltsch, Foraging personalities modify effects of habitat fragmentation on biodiversity, Oikos, 12 (2022), e09056. |
| [10] |
D. M. Debinski, R. D. Holt, A survey and overview of habitat fragmentation experiments, Conserv. Biol., 14 (2000), 342–355. https://doi.org/10.1046/j.1523-1739.2000.98081.x doi: 10.1046/j.1523-1739.2000.98081.x
|
| [11] |
A. Mai, G. Sun, F. Zhang, L. Wang, The joint impacts of dispersal delay and dispersal patterns on the stability of predator-prey metacommunities, J. Theor. Biol., 462 (2019), 455–465. https://doi.org/10.1016/j.jtbi.2018.11.035 doi: 10.1016/j.jtbi.2018.11.035
|
| [12] |
Y. Kang, S. K. Sasmal, K. Messan, A two-patch prey-predator model with predator dispersal driven by the predation strength, Math. Biosci. Eng., 14 (2017), 843–880. https://doi.org/10.3934/mbe.2017046 doi: 10.3934/mbe.2017046
|
| [13] |
J. Ban, Y. Wang, H. Wu, Dynamics of predator-prey systems with prey's dispersal between patches, Indian J. Pure Appl. Math., 53 (2022), 550–569. https://doi.org/10.1007/s13226-021-00117-5 doi: 10.1007/s13226-021-00117-5
|
| [14] |
K. Hu, Y. Wang, Dynamics of consumer-resource systems with consumer's dispersal between patches, Discrete Contin. Dyn. Syst. Ser. B, 27 (2022), 977–1000. https://doi.org/10.3934/dcdsb.2021077 doi: 10.3934/dcdsb.2021077
|
| [15] |
Z. Wang, Y. Wang, Bifurcations in diffusive predator–prey systems with Beddington–DeAngelis functional response, Nonlinear Dyn., 105 (2021), 1045–1061. https://doi.org/10.1007/s11071-021-06635-5 doi: 10.1007/s11071-021-06635-5
|
| [16] |
L. J. S. Allen, Persistence and extinction in single-species reaction-diffusion models, Bull. Math. Biol., 45 (1983), 209–227. https://doi.org/10.1016/S0092-8240(83)80052-4 doi: 10.1016/S0092-8240(83)80052-4
|
| [17] |
S. A. Levin, L. A. Segel, Hypothesis for origin of planktonic patchiness, Nature, 259 (1976), 659. https://doi.org/10.1038/259659a0 doi: 10.1038/259659a0
|
| [18] |
W. S. C. Gurney, R. M. Nisbet, The regulation of inhomogeneous populations, J. Theor. Biol., 52 (1975), 441–457. https://doi.org/10.1016/0022-5193(75)90011-9 doi: 10.1016/0022-5193(75)90011-9
|
| [19] |
X. Zhang, L. Chen, The linear and nonlinear diffusion of the competitive Lotka–Volterra model, Nonlinear Anal. Theory Methods Appl., 66 (2007), 2767–2776. https://doi.org/10.1016/j.na.2006.04.006 doi: 10.1016/j.na.2006.04.006
|
| [20] |
X. Zhou, X. Shi, X. Song, Analysis of nonautonomous predator-prey model with nonlinear diffusion and time delay, Appl. Math. Comput., 196 (2008), 129–136. https://doi.org/10.1016/j.amc.2007.05.041 doi: 10.1016/j.amc.2007.05.041
|
| [21] |
Z. Zhu, Y. Chen, Z. Li, F. Chen, Dynamic behaviors of a Leslie-Gower model with strong Allee effect and fear effect in prey, Math. Biosci. Eng., 20 (2023), 10977–10999. https://doi.org/10.3934/mbe.2023486 doi: 10.3934/mbe.2023486
|
| [22] |
Y. Liu, Z. Li, M. He, Bifurcation analysis in a Holling-Tanner predator-prey model with strong Allee effect, Math. Biosci. Eng., 20 (2023), 8632–8665. https://doi.org/10.3934/mbe.2023379 doi: 10.3934/mbe.2023379
|
| [23] |
T. Liu, L. Chen, F. Chen, Z. Li, Dynamics of a Leslie-Gower Model with weak Allee effect on prey and fear effect on predator, Int. J. Bifurcation Chaos, 33 (2023), 2350008. https://doi.org/10.1142/S0218127423500086 doi: 10.1142/S0218127423500086
|
| [24] |
T. Liu, L. Chen, F. Chen, Z. Li, Stability analysis of a Leslie-Gower model with strong Allee effect on prey and fear effect on predator, Int. J. Bifurcation Chaos, 32 (2022), 2250082. https://doi.org/10.1142/S0218127422500821 doi: 10.1142/S0218127422500821
|
| [25] |
Y. Lv, L. Chen, F. Chen, Z. Li, Stability and bifurcation in an SI epidemic model with additive Allee effect and time delay, Int. J. Bifurcation Chaos, 31 (2021), 2150060. https://doi.org/10.1142/S0218127421500607 doi: 10.1142/S0218127421500607
|
| [26] |
L. Chen, T. Liu, F. Chen, Stability and bifurcation in a two-patch model with additive Allee effect, AIMS Math., 7 (2022), 536–551. https://doi.org/10.3934/math.2022034 doi: 10.3934/math.2022034
|
| [27] |
W. Wang, Population dispersal and Allee effect, Ric. Mat., 65 (2016), 535–548. https://doi.org/10.1007/s11587-016-0273-0 doi: 10.1007/s11587-016-0273-0
|
| [28] |
H. Li, W. Yang, M. Wei, A. Wang, Dynamics in a diffusive predator–prey system with double Allee effect and modified Leslie–Gower scheme, Int. J. Biomath., 15 (2022), 2250001. https://doi.org/10.1142/S1793524522500012 doi: 10.1142/S1793524522500012
|
| [29] |
X. Hu, R. Sophia, The role of host refuge and strong Allee effects in a host–parasitoid system, Int. J. Biomath., 16 (2023), 2250107. https://doi.org/10.1142/S1793524522501078 doi: 10.1142/S1793524522501078
|
| [30] |
J. Geng, Y. Wang, Y. Liu, L. Yang, J. Yan, Analysis of an avian influenza model with Allee effect and stochasticity, Int. J. Biomath., 16 (2023), 2250111. https://doi.org/10.1142/S179352452250111X doi: 10.1142/S179352452250111X
|
| [31] |
X. Xu, Y. Meng, Y. Shao, Hopf bifurcation of a delayed predator–prey model with Allee effect and anti-predator behavior, Int. J. Biomath., 16 (2023), 2250125. https://doi.org/10.1142/S179352452250125X doi: 10.1142/S179352452250125X
|
| [32] |
J. B. Ferdy, J. Molofsky, Allee effect, spatial structure and species coexistence, J. Theor. Biol., 217 (2010), 3542–3556. https://doi.org/10.1016/j.amc.2010.09.029 doi: 10.1016/j.amc.2010.09.029
|
| [33] | Z. Zhang, T. Ding, W. Huang, Z. Dong, Qualitative Theory of Differential Equation, Science Press, Beijing, China. |
| [34] | L. Perko, Differential Equations and Dynamical Systems, Springer, New York, 1996. https://doi.org/10.1007/978-1-4684-0392-3 |
| [35] |
A. Dhooge, W. Govaerts, Y. A. Kuznetsov, MATCONT: A matlab package for numerical bifurcation analysis of odes, ACM Trans. Math. Software, 29 (2003), 141–164. https://doi.org/10.1145/779359.779362 doi: 10.1145/779359.779362
|
| [36] | D. Henry, Geometric Theory of Semilinear Parabolic Equations, Springer Berlin, Heidelberg, 2006. https://doi.org/10.1007/BFb0089647 |
| [37] |
V. Srivastava, E. M. Takyi, R. D. Parshad, The effect of fear on two species competition, Math. Biosci. Eng., 20 (2023), 8814–8855. https://doi.org/10.3934/mbe.2023388 doi: 10.3934/mbe.2023388
|
| [38] | S. Chen, F. Chen, V. Srivastava, R. D. Parshad, Dynamical analysis of a Lotka-Volterra competition model with both Allee and fear effect, Int. J. Biomath., (2023), forthcoming. |
| [39] |
G. Maciel, C. Cosner, R. B. Cantrell, F. Lutscher, Evolutionarily stable movement strategies in reaction–diffusion models with edge behavior, J. Math. Biol., 80 (2020), 61–92. https://doi.org/10.1007/s00285-019-01339-2 doi: 10.1007/s00285-019-01339-2
|
| [40] |
R. D. Parshad, E. Quansah, K. Black, M. Beauregard, Biological control via "ecological" damping: An approach that attenuates non-target effects, Math. Biosci., 273 (2016), 23–44. https://doi.org/10.1016/j.mbs.2015.12.010 doi: 10.1016/j.mbs.2015.12.010
|
| [41] | V. Srivastava, K. Antwi-Fordjour, R. D. Parshad, Exploring unique dynamics in a predator-prey model with generalist predator and group defense in prey, Chaos, 2023 (2023), forthcoming. |
Figures(13) / Tables(1)

Yue Xia, Lijuan Chen, Vaibhava Srivastava, Rana D. Parshad. Stability and bifurcation analysis of a two-patch model with the Allee effect and dispersal[J]. Mathematical Biosciences and Engineering, 2023, 20(11): 19781-19807. doi: 10.3934/mbe.2023876







 DownLoad:
DownLoad: