A delayed diffusive predator-prey system with nonmonotonic functional response subject to Neumann boundary conditions is introduced in this paper. First, we analyze the associated characteristic equation to research the conditions for local stability of the positive equilibrium point and the occurrence of Turing instability induced by diffusion in the absence of delay. Second, we provide conditions for the existence of Hopf bifurcation driven by time delay. By utilizing the normal theory and center manifold theorem, we derive explicit formulas for Hopf bifurcation properties such as direction and stability from the positive equilibrium. Third, a hybrid controller is added to the system. By judiciously adjusting the control parameters, we effectively enhance the stability domain of the system, resulting in a modification of the position of the Hopf bifurcation periodic solutions. Numerical simulations demonstrate the presence of rich dynamical phenomena within the system. Moreover, sensitivity analysis was conducted using Latin hypercube sampling (LHS)/partial rank correlation coefficient (PRCC) to explore the impact of parameter variations on the output of prey and predator populations.
Citation: Hairong Li, Yanling Tian, Ting Huang, Pinghua Yang. Hopf bifurcation and hybrid control of a delayed diffusive semi-ratio-dependent predator-prey model[J]. AIMS Mathematics, 2024, 9(10): 29608-29632. doi: 10.3934/math.20241434
A delayed diffusive predator-prey system with nonmonotonic functional response subject to Neumann boundary conditions is introduced in this paper. First, we analyze the associated characteristic equation to research the conditions for local stability of the positive equilibrium point and the occurrence of Turing instability induced by diffusion in the absence of delay. Second, we provide conditions for the existence of Hopf bifurcation driven by time delay. By utilizing the normal theory and center manifold theorem, we derive explicit formulas for Hopf bifurcation properties such as direction and stability from the positive equilibrium. Third, a hybrid controller is added to the system. By judiciously adjusting the control parameters, we effectively enhance the stability domain of the system, resulting in a modification of the position of the Hopf bifurcation periodic solutions. Numerical simulations demonstrate the presence of rich dynamical phenomena within the system. Moreover, sensitivity analysis was conducted using Latin hypercube sampling (LHS)/partial rank correlation coefficient (PRCC) to explore the impact of parameter variations on the output of prey and predator populations.

| [1] |
X. He, X. Zhao, T. Feng, Z. Qiu, Dynamical behaviors of a prey-predator model with foraging arena scheme in polluted environments, Math. Slovaca, 71 (2021), 235–250. http://doi.org/10.1515/ms-2017-0463. doi: 10.1515/ms-2017-0463

|
| [2] |
Y. Zhang, Q. Zhang, X. G. Yan, Complex dynamics in a singular Leslie-Gower predator-prey bioeconomic model with time delay and stochastic fluctuations, Phys. A: Stat. Mech. Appl., 404 (2014), 180–191. http://doi.org/10.1016/j.physa.2014.02.013 doi: 10.1016/j.physa.2014.02.013
|
| [3] | X. Q. Zhao, Dynamical systems in population biology, Berlin: Springer, 2003 http://doi.org/10.1007/978-0-387-21761-1 |
| [4] |
L. Zhang, C. Lu, Periodic solutions for a semi-ratio-dependent predator-prey system with Holling IV functional response, J. Appl. Math. Comput., 32 (2010), 465–477. https://doi.org/10.1007/s12190-009-0264-3 doi: 10.1007/s12190-009-0264-3
|
| [5] |
X. Xiu, A note on periodic solutions for semi-ratio-dependent predator-prey systems, Appl. Math. J. Chin. Univ., 25 (2010), 1–8. http://doi.org/10.1007/s11766-010-2106-3 doi: 10.1007/s11766-010-2106-3
|
| [6] |
B. Dai, Y. Li, Z. Luo, Multiple periodic solutions for impulsive Gause-type ratio-dependent predator-prey systems with non-monotonic numerical responses, Appl. Math. Comput., 217 (2011), 7478–7487. http://doi.org/10.1016/j.amc.2011.02.049 doi: 10.1016/j.amc.2011.02.049
|
| [7] | J. K. Zhuang, Periodicity for a semi-rati–dependent predator-prey system with delays on time scales, Int. J. Comput. Math. Sci., 4 (2010), 44–47. |
| [8] |
M. Zhao, Hopf bifurcation analysis for a semiratio-dependent predator-prey system with two delays, Abst. Appl. Anal., 9 (2013), 1140–1174. http://doi.org/10.1155/2013/495072 doi: 10.1155/2013/495072
|
| [9] |
J. Pradeesh, C. Vijayakumar, On the asymptotic stability of Hilfer fractional neutral stochastic differential systems with infinite delay, Qual. Theory Dyn. Syst., 23 (2024), 153. http://doi.org/10.1007/s12346-024-01007-x doi: 10.1007/s12346-024-01007-x
|
| [10] |
V. Gokulakrishnan, R. Srinivasan, Exponential input-to-state stabilization of stochastic nonlinear reaction-diffusion systems with time-varying delays and exogenous disturbances via boundary control, Comput. Appl. Math., 4 (2023), 308. http://dx.doi.org/10.1007/s40314-023-02447-y doi: 10.1007/s40314-023-02447-y
|
| [11] |
H. Achouri, C. Aouiti, Bogdanov-Takens and triple zero bifurcations for a neutral functional differential equations with multiple delays, J. Dyn. Diff. Equat., 35 (2023), 355–380. https://doi.org/10.1007/s10884-021-09992-2 doi: 10.1007/s10884-021-09992-2
|
| [12] |
A. Martin, S. Ruan, Predator-prey models with delay and prey harvesting, J. Math. Biol., 43 (2001), 247–267. https://doi.org/10.1007/s002850100095 doi: 10.1007/s002850100095
|
| [13] |
J. Wang, J. Wei, Bifurcation analysis of a delayed predator-prey system with strong Allee effect and diffusion, Appl. Anal., 91 (2012), 1219–1241. https://doi.org/10.1080/00036811.2011.563737 doi: 10.1080/00036811.2011.563737
|
| [14] |
R. Yuan, W. Jiang, Y. Wang, Saddle-node-Hopf bifurcation in a modified Leslie-Gower predator-prey model with time-delay and prey harvesting, J. Math. Anal. Appl., 422 (2015), 1072–1090. https://doi.org/10.1016/j.jmaa.2014.09.037 doi: 10.1016/j.jmaa.2014.09.037
|
| [15] |
Q. Chen, J. Gao, Hopf bifurcation and chaos control for a Leslie-Gower type generalist predator model, Adv. Differ. Equ., 2019 (2019), 315. https://doi.org/10.1186/s13662-019-2239-5 doi: 10.1186/s13662-019-2239-5
|
| [16] |
A. Kashkynbayev, A. Issakhanov, M. Otkel, J. Kurths, Finite-time and fixed-time synchronization analysis of shunting inhibitory memristive neural networks with time-varying delays, Chaos Solitons Fract., 156 (2022), 111866. https://doi.org/10.1016/j.chaos.2022.111866 doi: 10.1016/j.chaos.2022.111866
|
| [17] |
Y. Lv, L. Chen, F. Chen, Z. Li, Stability and bifurcation in an SI epidemic model with additive Allee effect and time delay, Int. J. Bifurc. Chaos, 31 (2021), 2150060. https://doi.org/10.1142/S0218127421500607 doi: 10.1142/S0218127421500607
|
| [18] |
S. Busenberg, W. Huang, Stability and Hopf bifurcation for a population delay model with diffusion effects, J. Differ. Equ., 124 (1996), 80–107. https://doi.org/10.1006/jdeq.1996.0003 doi: 10.1006/jdeq.1996.0003
|
| [19] |
A. M. Turing, The chemical basis of morphogenesisPhil, Trans. R. Soc. Lond. B, 237 (1952), 37–72. http://doi.org/10.1098/rstb.1952.0012 doi: 10.1098/rstb.1952.0012
|
| [20] |
V. V. Castets, E. Dulos, J. Boissonade, P. De Kepper, Experimental evidence of a sustained standing Turing-type nonequilibrium chemical pattern, Phys. Rev. Lett., 64 (1990), 2953–2956. https://doi.org/10.1103/PhysRevLett.64.2953 doi: 10.1103/PhysRevLett.64.2953
|
| [21] |
T. Y. Li, Q. R. Wang, Turing patterns in a predator-prey reaction-diffusion model with seasonality and fear effect, J. Nonlinear Sci., 33 (2023), 86. https://doi.org/10.1007/s00332-023-09938-6 doi: 10.1007/s00332-023-09938-6
|
| [22] |
P. Kumar, G. Gangopadhyay, Energetic and entropic cost due to overlapping of Turing-Hopf instabilities in the presence of cross diffusion, Phys. Rev. E, 101 (2020), 042204. https://doi.org/10.1103/PhysRevE.101.042204 doi: 10.1103/PhysRevE.101.042204
|
| [23] | J. Wu, Theory and applications of partial functional differential equations, Berlin: Springer, 1996. https://doi.org/10.1007/978-1-4612-4050-1 |
| [24] |
X. Zhang, H. Zhao, Bifurcation and optimal harvesting of a diffusive predator-prey system with delays and interval biological parameters, J. Theoret. Biol., 363 (2014), 390–403. https://doi.org/10.1016/j.jtbi.2014.08.031 doi: 10.1016/j.jtbi.2014.08.031
|
| [25] |
Y. Song, Q. Shi, Stability and bifurcation analysis in a diffusive predator-prey model with delay and spatial average, Math. Meth. Appl. Sci., 46 (2023), 5561–5584. http://doi.org/10.1002/mma.8853 doi: 10.1002/mma.8853
|
| [26] |
J. Liu, X. Zhang, Stability and Hopf bifurcation of a delayed reaction-diffusion predator-prey model with anti-predator behaviour, Nonlinear Anal.: Model. Control, 24 (2019), 387–406. http://doi.org/10.15388/NA.2019.3.5 doi: 10.15388/NA.2019.3.5
|
| [27] |
T. Wen, X. Wang, G. Zhang, Hopf bifurcation in a two-species reaction-diffusion-advection competitive model with nonlocal delay, Commun. Pure Appl. Anal., 22 (2023), 1517–1544. http://doi.org/10.3934/cpaa.2023036 doi: 10.3934/cpaa.2023036
|
| [28] | B. D. Hassard, N. D. Kazarinoff, Y. H. Wan, Theory and applications of Hopf bifurcation, Cambridge: Cambridge University Press, 1981. http://doi.org/10.1090/conm/445 |
| [29] |
Y. Qu, J. Wei, Bifurcation analysis in a time-delay model for prey-predator growth with stage-structure, Nonlinear Dyn., 49 (2007), 285–294. http://doi.org/10.1007/s11071-006-9133-x doi: 10.1007/s11071-006-9133-x
|
| [30] |
T. Y. Li, Q. R. Wang, Stability and Hopf bifurcation analysis for a two-species commensalism system with delay, Qual. Theory Dyn. Syst., 20 (2021), 83. https://doi.org/10.1007/s12346-021-00524-3 doi: 10.1007/s12346-021-00524-3
|
| [31] |
B. T. Mulugeta, L. Yu, Q. Yuan, J. Ren, Bifurcation analysis of a predator-prey model with strong Allee effect and Beddington-DeAngelis functional response, Discr. Cont. Dyn. Syst. B, 28 (2023), 1938–1963. http://doi.org/10.3934/dcdsb.2022153 doi: 10.3934/dcdsb.2022153
|
| [32] |
C. Huang, H. Li, J. Cao, A novel strategy of bifurcation control for a delayed fractional predator-prey model, Appl. Math. Comput., 347 (2019), 808–838. http://doi.org/10.1016/j.amc.2018.11.031 doi: 10.1016/j.amc.2018.11.031
|
| [33] |
S. Y. Li, Nonlinear delay-control of Hopf bifurcation and stability switches in a generlized logistic model, Annu. Int. Confer. Network Inform. Syst. Comput. (ICNISC), 8 (2022), 280–283. http://doi.org/10.1109/ICNISC57059.2022.00063 doi: 10.1109/ICNISC57059.2022.00063
|
| [34] |
T. Y. Li, Q. R. Wang, Bifurcation analysis for two-species commensalism (amensalism) systems with distributed delays, Int. J. Bifurc. Chaos Appl. Sci. Eng., 32 (2022), 2250133. http://doi.org/10.1142/S0218127422501334 doi: 10.1142/S0218127422501334
|
| [35] | X. W. Jiang, X. Y. Chen, T. W. Huang, H. C. Yan, Bifurcation and control for a predator-prey system with two delays, IEEE Trans. Circ. Syst. II: Express Briefs, 68 (2021), 376–380, |
| [36] |
A. Abta, H. Laarabi, T. Hamad, The Hopf bifurcation analysis and optimal control of a delayed SIR epidemic model, Int. J. Anal., 23 (2014), 1–10. http://doi.org/10.1155/2014/940819 doi: 10.1155/2014/940819
|
| [37] |
X. W. Jiang, X. Y. Chen, M. Chi, J. Chen, On Hopf bifurcation and control for a delay systems, Appl. Math. Comput., 370 (2020), 124906. https://doi.org/10.1016/j.amc.2019.124906 doi: 10.1016/j.amc.2019.124906
|
| [38] |
M. Xiao, G. Jiang, L. Zhao, State feedback control at Hopf bifurcation in an exponential RED algorithm model, Nonlinear Dyn., 76 (2014), 1469–1484. https://doi.org/10.1007/s11071-013-1221-0 doi: 10.1007/s11071-013-1221-0
|
| [39] |
W. Xu, T. Hayat, J. Cao, M. Xiao, Hopf bifurcation control for a fluid flow model of internet congestion control systems via state feedback, IMA J. Math. Control Inform., 33 (2016), 69–93. https://doi.org/10.1093/imamci/dnu029 doi: 10.1093/imamci/dnu029
|
| [40] |
X. S. Luo, G. Chen, B. H. Wang, J. Q. Fang, Hybrid control of period-doubling bifurcation and chaos in discrete nonlinear dynamical systems, Chaos Solitons Fract., 18 (2003), 775–783. https://doi.org/10.1016/s0960-0779(03)00028-6 doi: 10.1016/s0960-0779(03)00028-6
|
| [41] |
M. Peng, Z. Zhang, X. Wang, Hybrid control of Hopf bifurcation in a Lotka-Volterra predator-prey model with two delays, Adv. Diff. Equ., 1 (2017), 387. https://doi.org/10.1186/s13662-017-1434-5 doi: 10.1186/s13662-017-1434-5
|
| [42] |
P. Ghosh, Control of the Hopf-Turing transition by time-delayed global feedback in a reaction-diffusion system, Phys. Rev. E Stat. Nonlinear Soft Matter Phys., 84 (2011), 016222. https://doi.org/10.1103/PhysRevE.84.016222 doi: 10.1103/PhysRevE.84.016222
|
| [43] | R. R. Patra, S. Maitra, S. Kundu, Stability, bifurcation and control of a predator-prey ecosystem with prey herd behaviour against generalist predator with gestation delay, preprint paper, 2021. https://doi.org/10.48550/arXiv.2103.16263 |
| [44] |
R. R. Patra, S. Kundu, S. Maitra, Effect of delay and control on a predator-prey ecosystem with generalist predator and group defence in the prey species, Eur. Phys. J. Plus, 137 (2022), 128. https://doi.org/10.1140/epjp/s13360-021-02225-x doi: 10.1140/epjp/s13360-021-02225-x
|
| [45] |
B. Mondal, A. Sarkar, S. S. Santra, D. Majumder, T. Muhammad, Sensitivity of parameters and the impact of white noise on a generalist predator-prey model with hunting cooperation, Eur. Phys. J. Plus, 138 (2023), 1070. https://doi.org/10.1140/epjp/s13360-023-04710-x doi: 10.1140/epjp/s13360-023-04710-x
|
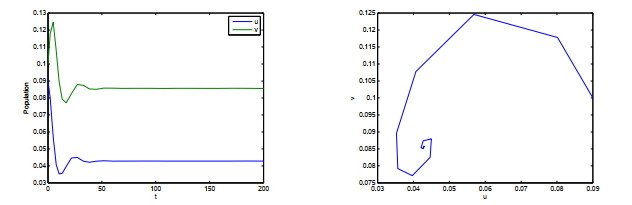
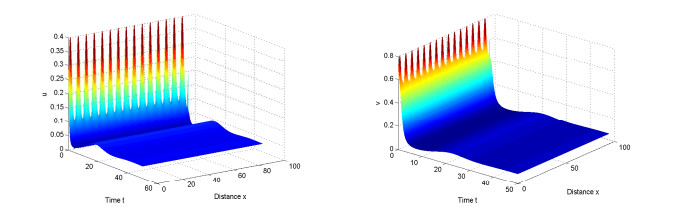
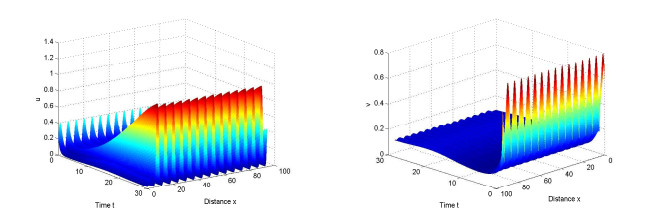
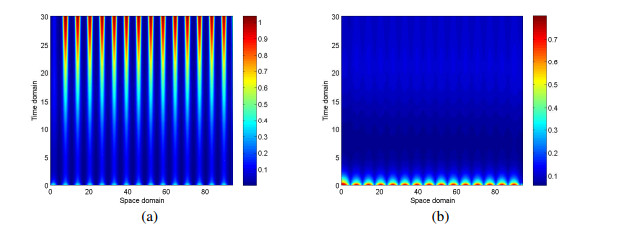
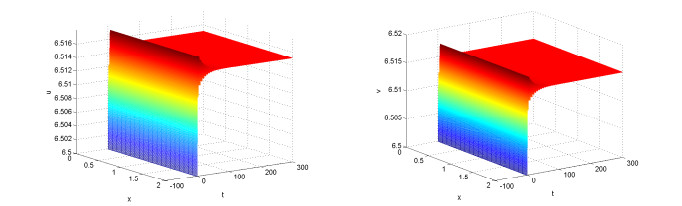
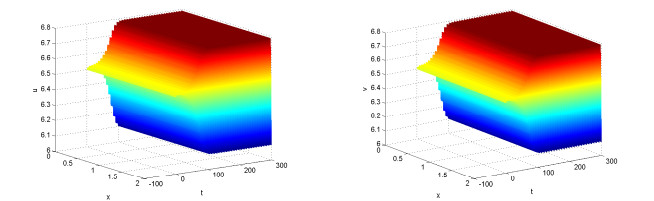
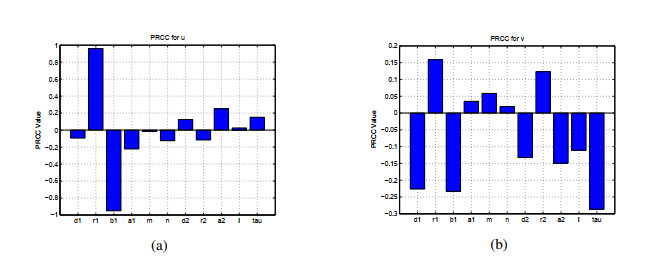
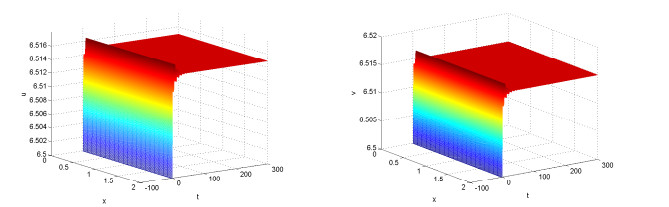
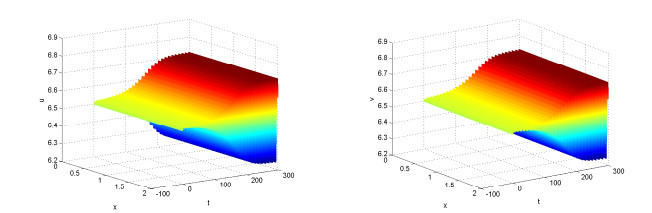
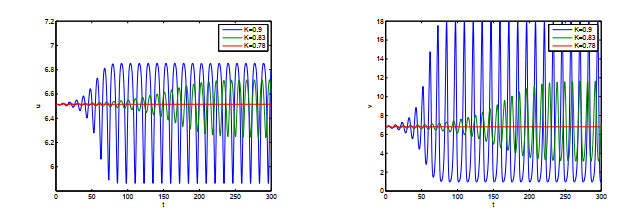
Figures(10)

Hairong Li, Yanling Tian, Ting Huang, Pinghua Yang. Hopf bifurcation and hybrid control of a delayed diffusive semi-ratio-dependent predator-prey model[J]. AIMS Mathematics, 2024, 9(10): 29608-29632. doi: 10.3934/math.20241434







 DownLoad:
DownLoad: