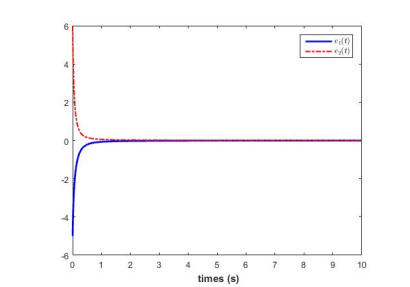
In this work, we study the fixed-time stability of fractional-order systems. By virtue of the properties of Riemann-Liouville fractional derivative and the comparison principle, we derive a new fixed-time stability theorem for fractional-order systems. Meanwhile, order-dependent setting time is formulated. Based on the developed fixed-time stability theorem, a fixed-time synchronization criterion for fractional-order neural networks is given. Simulation result demonstrates the effectiveness of our proposed results.
Citation: Yucai Ding, Hui Liu. A new fixed-time stability criterion for fractional-order systems[J]. AIMS Mathematics, 2022, 7(4): 6173-6181. doi: 10.3934/math.2022343
In this work, we study the fixed-time stability of fractional-order systems. By virtue of the properties of Riemann-Liouville fractional derivative and the comparison principle, we derive a new fixed-time stability theorem for fractional-order systems. Meanwhile, order-dependent setting time is formulated. Based on the developed fixed-time stability theorem, a fixed-time synchronization criterion for fractional-order neural networks is given. Simulation result demonstrates the effectiveness of our proposed results.

| [1] |
A. Polyakov, Nonlinear feedback design for fixed-time stabilization of linear control systems, IEEE Trans. Automat. Contr., 57 (2012), 2106–2110. http://dx.doi.org/10.1109/TAC.2011.2179869 doi: 10.1109/TAC.2011.2179869

|
| [2] |
J. D. Cao, R. X. Li, Fixed-time synchronization of delayed memristor-based recurrent neural networks, Sci. China Inf. Sci., 60 (2017), 032201. http://dx.doi.org/10.1007/s11432-016-0555-2 doi: 10.1007/s11432-016-0555-2
|
| [3] |
C. C. Hua, Y. F. Li, X. P. Guan, Finite/fixed-time stabilization for nonlinear interconnected systems with dead-zone input, IEEE Trans. Automat. Contr., 62 (2017), 2554–2560. http://dx.doi.org/10.1109/TAC.2016.2600343 doi: 10.1109/TAC.2016.2600343
|
| [4] |
C. Hu, J. Yu, Z. H. Chen, H. J. Jiang, T. W. Huang, Fixed-time stability of dynamical systems and fixed-time synchronization of coupled discontinuous neural networks, Neural Networks, 89 (2017), 74–83. http://dx.doi.org/10.1016/j.neunet.2017.02.001 doi: 10.1016/j.neunet.2017.02.001
|
| [5] |
F. Lopez-Ramirez, D. Efimov, A. Polyakov, W. Perruquetti, Conditions for fixed-time stability and stabilization of continuous autonomous systems, Syst. Control Lett., 129 (2019), 26–35. http://dx.doi.org/10.1016/j.sysconle.2019.05.003 doi: 10.1016/j.sysconle.2019.05.003
|
| [6] |
C. Chen, L. X. Li, H. P. Peng, Y. X. Yang, L. Mi, H. Zhao, A new fixed-time stability theorem and its application to the fixed-time synchronization of neural networks, Neural Networks, 123 (2020), 412–419. http://dx.doi.org/10.1016/j.neunet.2019.12.028 doi: 10.1016/j.neunet.2019.12.028
|
| [7] |
Q. Z. Xiao, H. L. Liu, X. Wang, Y. Huang, A note on the fixed-time bipartite flocking for nonlinear multi-agent systems, Appl. Math. Lett., 99 (2020), 105973. http://dx.doi.org/10.1016/j.aml.2019.07.004 doi: 10.1016/j.aml.2019.07.004
|
| [8] |
C. Aouiti, Q. Hui, H. Jallouli, E. Moulay, Fixed-time stabilization of fuzzy neutral-type inertial neural networks with time-varying delay, Fuzzy Sets Syst., 411 (2021), 48–67. http://dx.doi.org/10.1016/j.fss.2020.10.018 doi: 10.1016/j.fss.2020.10.018
|
| [9] | I. Podlubny, Geometric and physical interpretation of fractional integration and fractional differentiation, Journal of Fractional Calculus & Applied Analysis, 5 (2002), 367–386. |
| [10] | C. A. Monje, Y. Q. Chen, B. M. Vinagre, D. Y. Xue, V. Feliu-Batlle, Fractional-order systems and controls: fundamentals and applications, London: Springer, 2010. http://dx.doi.org/10.1007/978-1-84996-335-0 |
| [11] |
A. E. Matouk, A. A. Elsadany, Achieving synchronization between the fractional-order hyperchaotic Novel and Chen systems via a new nonlinear control technique, Appl. Math. Lett., 29 (2014), 30–35. http://dx.doi.org/10.1016/j.aml.2013.10.010 doi: 10.1016/j.aml.2013.10.010
|
| [12] |
F. F. Wang, D. Y. Chen, X. G. Zhang, Y. Wu, The existence and uniqueness theorem of the solution to a class of nonlinear fractional order system with time delay, Appl. Math. Lett., 53 (2016), 45–51. http://dx.doi.org/10.1016/j.aml.2015.10.001 doi: 10.1016/j.aml.2015.10.001
|
| [13] |
G. C. Wu, D. Baleanu, L. L. Huang, Novel Mittag-Leffler stability of linear fractional delay difference equations with impulse, Appl. Math. Lett., 82 (2018), 71–78. http://dx.doi.org/10.1016/j.aml.2018.02.004 doi: 10.1016/j.aml.2018.02.004
|
| [14] |
L. P. Chen, R. C. Wu, Y. Cheng, Y. Q. Chen, Delay-dependent and order-dependent stability and stabilization of fractional-order linear systems with time-varying delay, IEEE Trans. Circuits Syst. II, 67 (2020), 1064–1068. http://dx.doi.org/10.1109/TCSII.2019.2926135 doi: 10.1109/TCSII.2019.2926135
|
| [15] |
M. M. Li, J. R. Wang, Finite time stability of fractional delay differential equations, Appl. Math. Lett., 64 (2017), 170–176. http://dx.doi.org/10.1016/j.aml.2016.09.004 doi: 10.1016/j.aml.2016.09.004
|
| [16] |
X. Peng, H. Q. Wu, K. Song, J. X. Shi, Global synchronization in finite time for fractional-order neural networks with discontinuous activations and time delays, Neural Networks, 94 (2017), 46–54. http://dx.doi.org/10.1016/j.neunet.2017.06.011 doi: 10.1016/j.neunet.2017.06.011
|
| [17] |
V. N. Phat, N. T. Thanh, New criteria for finite-time stability of nonlinear fractional-order delay systems: A Gronwall inequality approach, Appl. Math. Lett., 83 (2018), 169–175. http://dx.doi.org/10.1016/j.aml.2018.03.023 doi: 10.1016/j.aml.2018.03.023
|
| [18] |
C. Rajivganthi, F. A. Rihan, S. Lakshmanan, P. Muthukumar, Finite-time stability analysis for fractional-order Cohen-Grossberg BAM neural networks with time delays, Neural Comput. & Applic., 29 (2018), 1309–1320. http://dx.doi.org/10.1007/s00521-016-2641-9 doi: 10.1007/s00521-016-2641-9
|
| [19] |
M. Syed Ali, G. Narayanan, Z. Orman, V. Shekher, S. Arik, Finite time stability analysis of fractional-order complex-valued memristive neural networks with proportional delays, Neural Process. Lett., 51 (2020), 407–426. http://dx.doi.org/10.1007/s11063-019-10097-7 doi: 10.1007/s11063-019-10097-7
|
| [20] |
S. Tyagi, S. C. Martha, Finite-time stability for a class of fractional-order fuzzy neural networks with proportional delay, Fuzzy Sets Syst., 381 (2020), 68–77. http://dx.doi.org/10.1016/j.fss.2019.04.010 doi: 10.1016/j.fss.2019.04.010
|
| [21] |
J. K. Ni, L. Liu, C. X. Liu, X. Y. Hu, Fractional order fixed-time nonsingular terminal sliding mode synchronization and control of fractional order chaotic systems, Nonlinear Dyn., 89 (2017), 2065–2083. http://dx.doi.org/10.1007/s11071-017-3570-6 doi: 10.1007/s11071-017-3570-6
|
| [22] |
Y. Sun, Y. Liu, Fixed-time synchronization of delayed fractional-order memristor-based fuzzy cellular neural networks, IEEE Access, 8 (2020), 165951–165962. http://dx.doi.org/10.1109/ACCESS.2020.3022928 doi: 10.1109/ACCESS.2020.3022928
|
| [23] |
P. Gong, Q. L. Han, Fixed-time bipartite consensus tracking of fractional-order multi-agent systems with a dynamic leader, IEEE Trans. Circuits Syst. II, 67 (2020), 2054–2058. http://dx.doi.org/10.1109/TCSII.2019.2947353 doi: 10.1109/TCSII.2019.2947353
|
| [24] |
M. Dutta, B. KrishnaRoy, A new memductance-based fractional-order chaotic system and its fixed-time synchronisation, Chaos Soliton. Fract., 145 (2021), 110782. http://dx.doi.org/10.1016/j.chaos.2021.110782 doi: 10.1016/j.chaos.2021.110782
|
| [25] |
M. Shirkavand, M. Pourgholi, Robust fixed-time synchronization of fractional order chaotic using free chattering nonsingular adaptive fractional sliding mode controller design, Chaos Soliton. Fract., 113 (2018), 135–147. http://dx.doi.org/10.1016/j.chaos.2018.05.020 doi: 10.1016/j.chaos.2018.05.020
|
| [26] |
S. Liu, X. Wu, X. F. Zhou, W. Jiang, Asymptotical stability of Riemann-Liouville fractional nonlinear systems, Nonlinear Dyn., 86 (2016), 65–71. http://dx.doi.org/10.1007/s11071-016-2872-4 doi: 10.1007/s11071-016-2872-4
|
| [27] |
C. Q. Long, G. D. Zhang, J. H. Hu, Fixed-time synchronization for delayed inertial complex-valued neural networks, Appl. Math. Comput., 405 (2021), 126272. http://dx.doi.org/10.1016/j.amc.2021.126272 doi: 10.1016/j.amc.2021.126272
|
| [28] |
G. Jumarie, Modified Riemann-Liouville derivative and fractional Taylor series of non-differentiable functions: Further results, Math. Comput. Appl., 51 (2006), 1367–1376. http://dx.doi.org/10.1016/j.camwa.2006.02.001 doi: 10.1016/j.camwa.2006.02.001
|
| [29] |
G. Jumarie, Informational entropy of non-random non-differentiable functions: an approach via fractional calculus, Appl. Math. Sci., 9 (2015), 2153–2185. http://dx.doi.org/10.12988/ams.2015.52139 doi: 10.12988/ams.2015.52139
|
| [30] |
P. D. Angelis, R. D. Marchis, A. L. Martire, I. Oliva, A mean-value approach to solve fractional differential and integral equations, Chaos Soliton. Fract., 138 (2020), 109895. http://dx.doi.org/10.1016/j.chaos.2020.109895 doi: 10.1016/j.chaos.2020.109895
|
| [31] | G. H. Hardy, J. E. Littlewood, G. Polya, Inequalities, Cambridge: Cambridge University Press, 1952. http://dx.doi.org/10.1017/S0025557200027455 |
| [32] |
Z. L. Wang, D. S. Yang, T. D. Ma, N. Sun, Stability analysis for nonlinear fractional-order systems based on comparison principle, Nonlinear Dyn., 75 (2014), 387–402. http://dx.doi.org/10.1007/s11071-013-1073-7 doi: 10.1007/s11071-013-1073-7
|

Figures(2)

Yucai Ding, Hui Liu. A new fixed-time stability criterion for fractional-order systems[J]. AIMS Mathematics, 2022, 7(4): 6173-6181. doi: 10.3934/math.2022343







 DownLoad:
DownLoad: