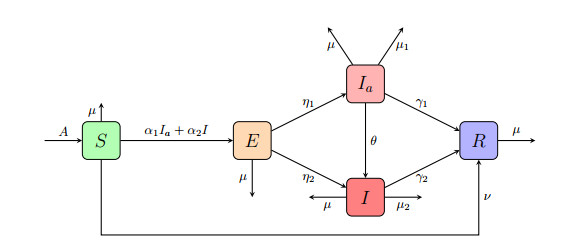
In this paper, we analyze the global dynamics of an endemic mathematical model that incorporates direct immunity by vaccination, as well as the shift from the asymptomatic to the symptomatic group in complex networks. By analyzing the Jacobian matrix and constructing suitable Lyapunov functionals, the stability of the disease-free equilibrium and the endemic equilibrium is determined with respect to the basic reproduction number $ R_0 $. Numerical simulations in scale-free and Poisson network environments are presented. The results validate the correctness of our theoretical analyses.
Citation: Erhui Li, Qingshan Zhang. Global dynamics of an endemic disease model with vaccination: Analysis of the asymptomatic and symptomatic groups in complex networks[J]. Electronic Research Archive, 2023, 31(10): 6481-6504. doi: 10.3934/era.2023328
In this paper, we analyze the global dynamics of an endemic mathematical model that incorporates direct immunity by vaccination, as well as the shift from the asymptomatic to the symptomatic group in complex networks. By analyzing the Jacobian matrix and constructing suitable Lyapunov functionals, the stability of the disease-free equilibrium and the endemic equilibrium is determined with respect to the basic reproduction number $ R_0 $. Numerical simulations in scale-free and Poisson network environments are presented. The results validate the correctness of our theoretical analyses.

| [1] |
Z. Ma, S. Wang, X. Lin, X. Li, X. Han, H. Wang, et al., Modeling for COVID-19 with the contacting distance, Nonlinear Dyn., 107 (2022), 3065–3084. https://doi.org/10.1007/s11071-021-07107-6 doi: 10.1007/s11071-021-07107-6

|
| [2] | J. Wieland, P. Mercorelli, Simulation of SARS-CoV-2 pandemic in Germany with ordinary differential equations in MATLAB, in 2021 25th International Conference on System Theory, Control and Computing (ICSTCC), (2021), 564–569. https://doi.org/10.1109/ICSTCC52150.2021.9607181 |
| [3] |
R. Yuan, Y. Ma, C. Shen, Z. Jinqing, X. Luo, M. Liu, Global dynamics of COVID-19 epidemic model with recessive infection and isolation, Math. Biosci. Eng., 18 (2021), 1833–1844. https://doi.org/10.3934/mbe.2021095 doi: 10.3934/mbe.2021095
|
| [4] |
H. Chen, B. Haus, P. Mercorelli, Extension of SEIR compartmental models for constructive Lyapunov control of COVID-19 and analysis in terms of practical stability, Mathematics, 9 (2021), 2227–7390. https://doi.org/10.3390/math9172076 doi: 10.3390/math9172076
|
| [5] |
P. Rakshit, S. Kumar, S. Noeiaghdam, U. Fernandez-Gamiz, M. Altanji, S. S. Santra, Modified SIR model for COVID-19 transmission dynamics: Simulation with case study of UK, US and India, Results Phys., 40 (2022), 105855. https://doi.org/10.1016/j.rinp.2022.105855 doi: 10.1016/j.rinp.2022.105855
|
| [6] | S. Guo, Y. Xue, X. Li, Z. Zheng, Dynamics of COVID-19 models with asymptomatic infections and quarantine measures, preprint, (2022). https://doi.org/10.21203/rs.3.rs-2291574/v1 |
| [7] |
S. H. Khoshnaw, M. Shahzad, M. Ali, F. Sultan, A quantitative and qualitative analysis of the COVID–19 pandemic model, Chaos Solitons Fractals, 138 (2020), 109932. https://doi.org/10.1016/j.chaos.2020.109932 doi: 10.1016/j.chaos.2020.109932
|
| [8] | J. B. Aguilar, J. S. Faust, L. M. Westafer, J. B. Gutierrez, A model describing COVID-19 community transmission taking into account asymptomatic carriers and risk mitigation, medRxiv preprint, (2020). https://doi.org/10.1101/2020.03.18.20037994 |
| [9] |
M. Peirlinck, K. Linka, F. Sahli Costabal, J. Bhattacharya, E. Bendavid, J. P. Ioannidis, et al., Visualizing the invisible: {T}he effect of asymptomatic transmission on the outbreak dynamics of COVID-19, Comput. Methods Appl. Mech. Eng., 372 (2020), 113410. https://doi.org/10.1016/j.cma.2020.113410 doi: 10.1016/j.cma.2020.113410
|
| [10] |
S. M. A. Rahman, X. Zou, Flu epidemics: a two-strain flu model with a single vaccination, J. Biol. Dyn., 5 (2011), 376–390. https://doi.org/10.1080/17513758.2010.510213 doi: 10.1080/17513758.2010.510213
|
| [11] |
M. Y. Li, Z. Shuai, C. Wang, Global stability of multi-group epidemic models with distributed delays, J. Math. Anal. Appl., 361 (2010), 38–47. https://doi.org/10.1016/j.jmaa.2009.09.017 doi: 10.1016/j.jmaa.2009.09.017
|
| [12] |
I. A. Baba, B. Kaymakamzade, E. Hincal, Two-strain epidemic model with two vaccinations, Chaos Solitons Fractals, 106 (2018), 342–348. https://doi.org/10.1016/j.chaos.2017.11.035 doi: 10.1016/j.chaos.2017.11.035
|
| [13] |
B. Kaymakamzade, E. Hincal, Two-strain epidemic model with two vaccinations and two time delayed, Qual. Quant., 52 (2018), 695–709. https://doi.org/10.1007/s11135-017-0647-8 doi: 10.1007/s11135-017-0647-8
|
| [14] |
M. Fudolig, R. Howard, The local stability of a modified multi-strain SIR, PLOS ONE, 15 (2020), 1–27. https://doi.org/10.1371/journal.pone.0243408 doi: 10.1371/journal.pone.0243408
|
| [15] |
S. Huang, F. Chen, L. Chen, Global dynamics of a network-based SIQRS epidemic model with demographics and vaccination, Commun. Nonlinear Sci. Numer. Simul., 43 (2017), 296–310. https://doi.org/10.1016/j.cnsns.2016.07.014 doi: 10.1016/j.cnsns.2016.07.014
|
| [16] |
M. Sun, H. Zhang, H. Kang, Epidemic spreading on adaptively weighted scale-free networks, J. Math. Biol., 74 (2017), 1263–1298. https://doi.org/10.1007/s00285-016-1057-6 doi: 10.1007/s00285-016-1057-6
|
| [17] |
X. Meng, Z. Cai, S. Si, D. Duan, Analysis of epidemic vaccination strategies on heterogeneous networks: Based on SEIRV model and evolutionary game, Appl. Math. Comput., 403 (2021), 126172. https://doi.org/10.1016/j.amc.2021.126172 doi: 10.1016/j.amc.2021.126172
|
| [18] |
W. Lv, Q. Ke, K. Li, Dynamical analysis and control strategies of an SIVS epidemic model with imperfect vaccination on scale-free networks, Nonlinear Dyn., 99 (2020), 1507–1523. https://doi.org/10.1007/s11071-019-05371-1 doi: 10.1007/s11071-019-05371-1
|
| [19] |
C. H. Li, A. M. Yousef, Bifurcation analysis of a network-based SIR epidemic model with saturated treatment function, Chaos Interdiscip. J. Nonlinear Sci., 29 (2019), 033129. https://doi.org/10.1063/1.5079631 doi: 10.1063/1.5079631
|
| [20] |
Y. Yao, J. Zhang, A two-strain epidemic model on complex networks with demographics, J. Biol. Syst., 24 (2016), 577–609. https://doi.org/10.1142/S0218339016500297 doi: 10.1142/S0218339016500297
|
| [21] |
J. Yang, C. H. Li, Dynamics of a competing two-strain SIS epidemic model on complex networks with a saturating incidence rate, J. Phys. A: Math. Theor., 49 (2016), 215601. https://doi.org/10.1088/1751-8113/49/21/215601 doi: 10.1088/1751-8113/49/21/215601
|
| [22] |
X. Cheng, Y. Wang, G. Huang, Global dynamics of a network-based SIQS epidemic model with nonmonotone incidence rate, Chaos Solitons Fractals, 153 (2021), 111502. https://doi.org/10.1016/j.chaos.2021.111502 doi: 10.1016/j.chaos.2021.111502
|
| [23] |
J. Wang, M. Liu, Y. Li, Global stability analysis of an SIR epidemic model with demographics and time delay on networks, Physica A: Stat. Mech. Appl., 410 (2014), 268–275. https://doi.org/10.1016/j.physa.2014.05.011 doi: 10.1016/j.physa.2014.05.011
|
| [24] |
P. Yang, Y. Wang, Dynamics for an SEIRS epidemic model with time delay on a scale-free network, Physica A: Stat. Mech. Appl., 527 (2019), 121290. https://doi.org/10.1016/j.physa.2019.121290 doi: 10.1016/j.physa.2019.121290
|
| [25] |
R. Pastor-Satorras, A. Vespignani, Epidemic spreading in scale-free networks, Phys. Rev. Lett., 86 (2001), 3200–3203. https://doi.org/10.1103/PhysRevLett.86.3200 doi: 10.1103/PhysRevLett.86.3200
|
| [26] | J. P. La Salle, The Stability of Dynamical Systems, Society for Industrial and Applied Mathematics, Philadelphia, 1994. http://dx.doi.org/10.1137/1.9781611970432 |
| [27] |
M. Y. Li, Z. Shuai, Global-stability problem for coupled systems of differential equations on networks, J. Differ. Equations, 248 (2010), 1–20. https://doi.org/10.1016/j.jde.2009.09.003 doi: 10.1016/j.jde.2009.09.003
|
| [28] | P. Erdős, A. Rényi, On random graphs I, Publ. Math. Debrecen, 6 (1959), 290–297. |
| [29] |
P. H. T. Schimit, F. H. Pereira, Disease spreading in complex networks: A numerical study with Principal Component Analysis, Expert Syst. Appl., 97 (2018), 41–50. https://doi.org/10.1016/j.eswa.2017.12.021 doi: 10.1016/j.eswa.2017.12.021
|
| [30] |
A. L. Barabási, R. Albert, Emergence of scaling in random networks, Science, 286 (1999), 509–512. https://doi.org/10.1126/science.286.5439.509 doi: 10.1126/science.286.5439.509
|









Figures(10) / Tables(1)
Erhui Li, Qingshan Zhang. Global dynamics of an endemic disease model with vaccination: Analysis of the asymptomatic and symptomatic groups in complex networks[J]. Electronic Research Archive, 2023, 31(10): 6481-6504. doi: 10.3934/era.2023328







 DownLoad:
DownLoad: