Citation: Gerardo Aquino, Andrea Rocco. Bimodality in gene expression without feedback: from Gaussian white noise to log-normal coloured noise[J]. Mathematical Biosciences and Engineering, 2020, 17(6): 6993-7017. doi: 10.3934/mbe.2020361

| [1] |
J. R. Pomerening, Uncovering mechanisms of bistability in biological systems, Curr. Opin. Biotechnol., 19 (2008), 381-388. doi: 10.1016/j.copbio.2008.06.009

|
| [2] |
M. C. A. Leite, Y. Wang, Multistability, oscillations and bifurcations in feedback loops, Math. Biosci. Eng., 7 (2010), 83-97. doi: 10.3934/mbe.2010.7.83
|
| [3] |
G. Balázsi, A. van Oudenaarden, J. J. Collins, Cellular decision-making and biological noise: From microbes to mammals, Cell, 144 (2011), 910-925. doi: 10.1016/j.cell.2011.01.030
|
| [4] |
L. Wang, M. C. Romano, F. A. Davidson, Translational control of gene expression via interacting feedback loops, Phys. Rev. E, 100 (2019), 050402. doi: 10.1103/PhysRevE.100.050402
|
| [5] |
M. Santillan, M. C. Mackey, E. S. Zeron, Origin of bistability in the lac operon, Biophys. J., 92 (2007), 3830-3842. doi: 10.1529/biophysj.106.101717
|
| [6] |
J. W. Williams, X. Cui, A. Levchenko, A. M. Stevens, Robust and sensitive control of a quorum-sensing circuit by two interlocked feedback loops, Mol. Sys. Biol., 4 (2008), 234. doi: 10.1038/msb.2008.70
|
| [7] |
P. Melke, P. Sahlin, A. Levchenko, H. Jonsson, A cell-based model for quorum sensing in heterogeneous bacterial colonies, PLoS Comp. Biol., 6 (2010), e1000819. doi: 10.1371/journal.pcbi.1000819
|
| [8] | J. E. Ferrell, Bistability, bifurcations, and Waddington's epigenetic landscape, Curr. Biol., 22 (2012), R458-R466. |
| [9] |
M. B. Elowitz, A. J. Levine, E. D. Siggia, P. S. Swain, Stochastic gene expression in a single cell, Science, 297 (2002), 1183-1186. doi: 10.1126/science.1070919
|
| [10] | A. P. Gasch, F. B. Yu, J. Hose, L. E. Escalante, M. Place, R. Bacher, et al., Single-cell RNA sequencing reveals intrinsic and extrinsic regulatory heterogeneity in yeast responding to stress, PLOS Biol., 15 (2017), e2004050. |
| [11] | H. Ochiai, T. Hayashi, M. Umeda, M. Yoshimura, A. Harada, Y. Shimizu, et al., Genome-wide analysis of transcriptional bursting-induced noise in mammalian cells, preprint, BioRxiv, 2019: 736207. |
| [12] | J. M. Raser, E. K. O'Shea, Noise in gene expression: Origins, consequences, and control, Science, 309 (2005), 2010-2013. |
| [13] |
A. Eldar, M. B. Elowitz, Functional roles for noise in genetic circuits, Nature, 467 (2010), 167-173. doi: 10.1038/nature09326
|
| [14] | L. S. Tsimring, Noise in biology, Rep. Prog. Phys., 77 (2014), 026601. |
| [15] | N. G. Van Kampen, Stochastic Processes in Physics and Chemistry, North-Holland, 2007. |
| [16] |
J. Elf, M. Ehrenberg, Fast evaluation of fluctuations in biochemical networks with the linear noise approximation, Genome Res., 13 (2003), 2475-2484. doi: 10.1101/gr.1196503
|
| [17] |
D. T. Gillespie, Exact stochastic simulations of coupled chemical reactions, J. Phys. Chem., 81 (1977), 2340-2361. doi: 10.1021/j100540a008
|
| [18] |
D. T. Gillespie, Approximate accelerated stochastic simulation of chemically reacting systems, J. Chem. Phys., 115 (2001), 1716-1733. doi: 10.1063/1.1378322
|
| [19] |
P. S. Swain, M. B. Elowitz, E. D. Siggia, Intrinsic and extrinsic contributions to stochasticity in gene expression, Proc. Natl. Acad. Sci., 99 (2002), 12795-12800. doi: 10.1073/pnas.162041399
|
| [20] |
J. Dattani, M. Barahona, Stochastic models of gene transcription with upstream drives: Exact solution and sample path characterisation, J. R. Soc. Interface, 14 (2017), 20160833. doi: 10.1098/rsif.2016.0833
|
| [21] |
P. C. Bressloff, E. Levien, Propagation of extrinsic fluctuations in biochemical birth-death processes, Bull. Math. Biol., 81 (2019), 800-829. doi: 10.1007/s11538-018-00538-0
|
| [22] | W. Horsthemke, R. Lefever, Noise-Induced Transitions, Springer, 1984. |
| [23] | C. W. Gardiner, Handbook of Stochastic Methods, Springer, 1985. |
| [24] | J. Garcia-Ojalvo, J. Sancho, Noise in Spatially Extended Systems, Springer, 1999. |
| [25] |
C. Zeng, C. Zhang, J. Zeng, H. Luo, D. Tian, H. Zhang, et al., Noises-induced regime shifts and -enhanced stability under a model of lake approaching eutrophication, Ecol. Complexity, 22 (2015), 102-108. doi: 10.1016/j.ecocom.2015.02.005
|
| [26] |
J. Jungeilgesa, T. Ryazanova, Noise-induced transitions in a stochastic Goodwin-type business cycle model, Struct. Change and Econ. Dyn. 40 (2017), 103-115. doi: 10.1016/j.strueco.2017.01.003
|
| [27] |
M. Samoilov, S. Plyasunov, A. P. Arkin, Stochastic amplification and signaling in enzymatic futile cycles through noise-induced bistability with oscillations, Proc. Natl. Acad. Sci., 102 (2005), 2310-2315. doi: 10.1073/pnas.0406841102
|
| [28] |
M. Samoilov, A. P. Arkin, Deviant effects in molecular reaction pathways, Nat. Biotechnol., 24 (2006), 1235-1240. doi: 10.1038/nbt1253
|
| [29] |
A. Rocco, Stochastic control of metabolic pathways, Phys. Biol., 6 (2009), 016002. doi: 10.1088/1478-3975/6/1/016002
|
| [30] |
E. Pujadas, A. P. Feinberg, A role for regulated noise in the epigenetic landscape of development and disease, Cell, 148 (2012), 1123-1131. doi: 10.1016/j.cell.2012.02.045
|
| [31] |
M. Weber, J. Buceta, Dynamics of the quorum sensing switch: stochastic and non-stationary effects, BMC Syst. Biol., 7 (2013), 6. doi: 10.1186/1752-0509-7-6
|
| [32] |
M. Weber, J. Buceta, Stochastic stabilization of phenotypic states: The genetic bistable switch as a case study, PLOS One, 8 (2013), e73487. doi: 10.1371/journal.pone.0073487
|
| [33] | A. Ochab-Marcinek, M. Tabaka, Bimodal gene expression in noncooperative regulatory systems Proc. Natl. Acad. Sci., 107 (2010), 22096-22101. |
| [34] |
A. Ochab-Marcinek, J. Jedrak, M. Tabaka, Hill Kinetics as a noise filter: The role of transcription factor autoregulation in gene cascades, Phys. Chem. Chem. Phys., 19 (2017), 22580-22591. doi: 10.1039/C7CP00743D
|
| [35] |
M. R. Birtwistle, J. Rauch, A. Kiyatkin, E. Aksamitiene, M. Dobrzyński, J. B. Hoek, et al., Emergence of bimodal cell population responses from the interplay between analog single-cell signaling and protein expression noise, BMC Syst. Biol., 6 (2012), 109. doi: 10.1186/1752-0509-6-109
|
| [36] |
K. H. Kim, H. M. Sauro, In search of noise-induced bimodality, BMC Biol., 10 (2012), 89. doi: 10.1186/1741-7007-10-89
|
| [37] |
M. Dobrzyński, L. K. Nguyen, M. R. Birtwistle, A. von Kriegsheim, A. B. Fernàndez, A. Cheong, et al., Nonlinear signalling networks and cell- to-cell variability transform external signals into broadly distributed or bimodal responses, J. R. Soc. Interface, 11 (2014), 20140383. doi: 10.1098/rsif.2014.0383
|
| [38] |
N. Q. Balaban, J. Merrin, R. Chait, L. Kowalik, S. Leibler, Bacterial persistence as a phenotypic switch, Science, 305 (2004), 1622-1625. doi: 10.1126/science.1099390
|
| [39] | C. Lou, Z. Li, Q. Ouyang, A molecular model for persister in E. coli, J. Theo. Biol., 255 (2008), 205-209 |
| [40] | S. M. Hingley-Wilson, N. Ma, Y. Hu, R. Casey, A. Bramming, R. J. Curry, et al., Loss of phenotypic inheritance associated with ydcI mutation leads to increased frequency of small, slow persisters in escherichia coli, Proc. Natl. Acad. Sci., 117 (2020), 4152-4157. |
| [41] |
A. Rocco, A. Kierzek, J. McFadden, Slow protein fluctuations explain the emergence of growth phenotypes and persistence in clonal bacterial populations, PloS One, 8 (2013), e54272. doi: 10.1371/journal.pone.0054272
|
| [42] | A. Rocco, A. Kierzek, J. McFadden, Systems Biology of Tuberculosis, Springer, 2013. |
| [43] | L. Arnold, W. Horsthemke, R. Lefever, White and coloured external noise and transition phenomena in nonlinear systems, Z. Phys. B Condens. Matter, 29 (1978), 367-373. |
| [44] | L. E. Reichl, W. C. Schieve, Instabilities, Bifurcation, and Fluctuations in Chemical Systems, University of Texas Press, Austin, 1982. |
| [45] |
N. Rosenfeld, J. W. Young, U. Alon, P. S. Swain, M. B. Elowitz, Gene regulation at the single-cell level, Science, 307 (2005), 1962-1965. doi: 10.1126/science.1106914
|
| [46] | B. B. Kaufmann, Q. Yang, J. T. Mettetal, A. van Oudenaarden, Heritable stochastic switching revealed by single-cell genealogy, PLoS Biol., 5 (2007), 1973-1980. |
| [47] |
P. Jung, P. Hänggi, Dynamical systems: A unified colored-noise approximation, Phys. Rev. A, 35 (1987), 4464-4466. doi: 10.1103/PhysRevA.35.4464
|
| [48] |
P. Grigolini, L. A. Lugiato, R. Mannella, P. V. E. McClintock, M. Merri, M. Pernigo, Fokker-pLanck description of stochastic processes with colored noise, Phys Rev. A, 38 (1988), 1966. doi: 10.1103/PhysRevA.38.1966
|
| [49] | J. Holehouse, A. Gupta, R. Grima, Steady-state fluctuations of a genetic feedback loop with fluctuating rate parameters using the unified colored noise approximation, J. Phys. A: Math. Theor., Forthcoming. |
| [50] |
L. Borland, Itô-Langevin equations within generalized thermostatistics, Phys. Lett. A, 245 (1998), 67-72. doi: 10.1016/S0375-9601(98)00467-8
|
| [51] |
R. V. Bobryk, A. Chrzeszczyk, Transitions induced by bounded noise, Phys. A, 358 (2005), 263-272. doi: 10.1016/j.physa.2005.03.055
|
| [52] | S. de Franciscis, G. Caravagna, A. d'Onofrio, Bounded noises as a natural tool to model extrinsic fluctuations in biomolecular networks, Nat. Comput. 13 (2014), 297-307. |
| [53] |
H. S. Wio, R. Toral, Effect of non-Gaussian noise sources in a noise induced transition, Phys. D, 193 (2004), 161-168. doi: 10.1016/j.physd.2004.01.017
|
| [54] | A. d'Onofrio, Bounded-noise-induced transitions in a tumor-immune system interplay, Phys. Rev. E, 81 (2010), 021923. |
| [55] |
A. d'Onofrio, A. Gandolfi, Resistance to antitumor chemotherapy due to bounded-noise-induced transitions, Phys. Rev. E, 82 (2010), 061901. doi: 10.1103/PhysRevE.82.061901
|
| [56] |
L. Cai, N. Friedman, X. S. Xie, Stochastic protein expression in individual cells at the single molecule level, Nature, 440 (2006), 358-362. doi: 10.1038/nature04599
|
| [57] | M. Bengtsson, A. Stahlberg, P. Rorsman, M. Kubista, Gene expression profiling in single cells from the pancreatic islets of langerhans reveals lognormal distribution of mRNA levels, Genome Res., 5 (2005), 1388-1392. |
| [58] |
L. Ham, R. D. Brackston, M. P. H. Stumpf, Extrinsic noise and heavy-tailed laws in gene expression, Phys. Rev. Lett., 124 (2020), 108101. doi: 10.1103/PhysRevLett.124.108101
|
| [59] |
D. T. Gillespie, A rigorous derivation of the chemical master equation, Phys. A, 188 (1992), 404-425. doi: 10.1016/0378-4371(92)90283-V
|
| [60] |
D. T. Gillespie, The chemical Lengevin equation, J. Chem. Phys., 113 (2000), 297. doi: 10.1063/1.481811
|
| [61] |
F. Wong, M. Zakai, On the convergence of ordinary integrals to stochastic integrals, Ann. Math. Stat., 36 (1965), 1560-1564. doi: 10.1214/aoms/1177699916
|
| [62] |
V. Shahrezaei, J. F. Ollivier, P. S. Swain, Colored extrinsic fluctuations and stochastic gene expression, Mol. Sys. Bio., 4 (2008), 196. doi: 10.1038/msb.2008.31
|
| [63] |
N. Rosenfeld, M. B. Elowitz, U. Alon, Negative autoregulation speeds the response times of transcription networks, J. Mol. Biol., 323 (2002), 785-793. doi: 10.1016/S0022-2836(02)00994-4
|
| [64] | M Thattai, A. van Oudenaarden, Intrinsic noise in gene regulatory networks, Proc. Natl. Acad. Sci., 98 (2001), 8614. |
| [65] | B. H. Toyama, M. W. Hetzer, Protein homeostasis: Live long, won't prosper, Nat. Rev. Mol. Cell. Biol., 14 (2013), 55-61. |
| [66] | A. Loinger, A. Lipshtat, N. Q. Balaban, O. Biham, Stochastic simulations of genetic switch systems, Phys Rev., 75 (2007), 021904. |
| [67] |
G. Q. Cai, Y. K. Lin, Generation of non-Gaussian stationary stochastic processes, Phys. Rev. E, 54 (1996), 299-303. doi: 10.1103/PhysRevE.54.299
|
| [68] |
M. Kessler, M. Sørensen, Estimating equations based on eigenfunctions for a discretely observed diffusion process, Bernoulli, 5 (1999), 299-314. doi: 10.2307/3318437
|
| [69] |
N. Friedman, L. Cai, X. S. Xie, Linking stochastic dynamics to population distribution: An analytical framework of gene expression, Phys. Rev. Lett., 97 (2006), 168302. doi: 10.1103/PhysRevLett.97.168302
|
| [70] |
V. Shahrezaei, P. S. Swain, Analytical distributions for stochastic gene expression, Proc. Natl. Acad. Sci., 105 (2008), 17256-17261. doi: 10.1073/pnas.0803850105
|
| [71] | Y. Taniguchi, P. J. Choi, G. W. Li, H. Chen, M. Babu, J. Hearn, et al., Quantifying E. coli proteome and transcriptome with single-molecule sensitivity in single cells, Science, 329 (2010), 533-538. |
| [72] |
H. Salman, N. Brenner, C. K. Tung, N. Elyahu, E. Stolovicki, L. Moore, et al., Universal protein fluctuations in populations of microorganisms, Phys. Rev. Lett., 108 (2012), 238105. doi: 10.1103/PhysRevLett.108.238105
|
| [73] |
N. Brenner, C. M. Newman, D. Osmanovic, Y. Rabin, H. Salman, D. L. Stein, Universal protein distributions in a model of cell growth and division, Phys. Rev. E, 92 (2015), 042713. doi: 10.1103/PhysRevE.92.042713
|
| [74] |
A. Sigal, R. Milo, A. Cohen, N. Geva-Zatorsky, Y. Klein, Y. Liron, et al., Variability and memory of protein levels in human cells, Nature, 444 (2006), 643-646. doi: 10.1038/nature05316
|
| [75] |
J. Lei, Stochasticity in single gene expression with both intrinsic noise and fluctuation in kinetic parameters, J. Theo. Biol., 256 (2009), 485-492. doi: 10.1016/j.jtbi.2008.10.028
|
| [76] |
F. Maleki, A. Becskei, An open-loop approach to calculate noise-induced transitions, J. Theo. Biol., 415 (2017), 145-157. doi: 10.1016/j.jtbi.2016.12.012
|
| [77] | D. Angeli, J. E. Ferrell Jr, E.D. Sontag, Detection of multistability, bifurcations, and hysteresis in a large class of biological positive-feedback systems, Proc. Natl. Acad.. Sci., 101 (2004), 1822-1827. |
| [78] |
C. Hsu, V. Jaquet, F. Maleki, A. Becskei, Contribution of bistability and noise to cell fate tansitions determined by feedback opening, J. Mol. Biol., 428 (2016), 4115-4128. doi: 10.1016/j.jmb.2016.07.024
|
| [79] |
R. Kupferman, G. A. Pavliotis, A. M. Stuart, Itô versus Stratonovich white noise limits for systems with inertia and colored multiplicative noise, Phys. Rev. E, 70 (2004), 036120. doi: 10.1103/PhysRevE.70.036120
|
| [80] |
G. A. Pavliotis, A. M. Stuart, Analysis of white noise limits for stochastic systems with two fast relaxation times, Multiscale Model. Simul., 4 (2005), 1. doi: 10.1137/040610507
|
| [81] | P. Thomas, R. Grima, A. V. Straube, Rigorous elimination of fast stochastic variables from the linear noise approximation using projection operators, Phys. Rev. E, 86 (2012), 041110. |
| [82] | B. Bravi, K. J. Rubin, P. Sollich, Systematic model reduction captures the dynamics of extrinsic noise inbiochemical subnetworks, preprint, arXiv: 2003.08704. |
| [83] |
R. Perez-Carrasco, P. Guerrero, J. Briscoe, K. M. Page, Intrinsic noise profoundly alters the dynamics and steady state of morphogen-controlled bistable genetic switches, PLoS Comput. Biol., 12 (2016), e1005154. doi: 10.1371/journal.pcbi.1005154
|
| [84] |
M. Pájaro, O. Otero-Muras, C. Vázquez, A. A. Alonso, Transient hysteresis and inherent stochasticity in gene regulatroy networks, Nat. Commun., 10 (2019), 4581. doi: 10.1038/s41467-019-12344-w
|
| [85] |
A. Brock, H. Chang, S. Huang, Non-genetic heterogeneity—a mutation—independent driving force for the somatic evolution of tumours, Nat. Rev. Gen., 10 (2009), 336-342. doi: 10.1038/nrg2556
|
| [86] |
A Sanchez, S. Choubey, J. Kondev, Regulation of noise in gene expression, Annu. Rev. Biophys., 42 (2013), 469-491. doi: 10.1146/annurev-biophys-083012-130401
|
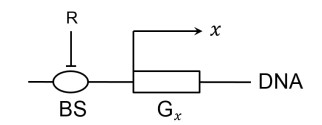
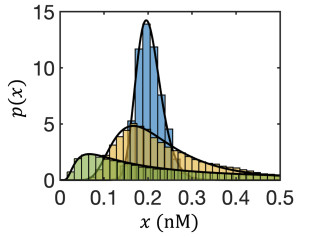
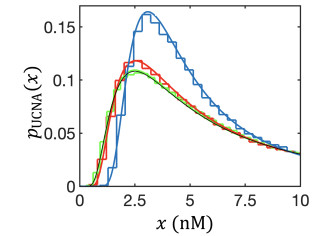
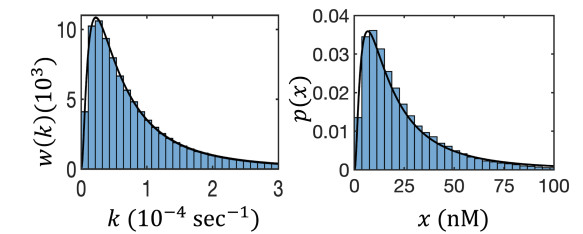
Figures(8)

Gerardo Aquino, Andrea Rocco. Bimodality in gene expression without feedback: from Gaussian white noise to log-normal coloured noise[J]. Mathematical Biosciences and Engineering, 2020, 17(6): 6993-7017. doi: 10.3934/mbe.2020361







 DownLoad:
DownLoad: