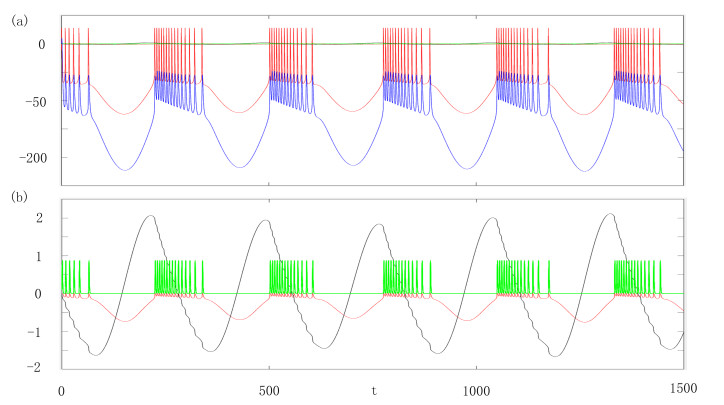
Half-center oscillators are typical small circuits that are crucial for understanding CPG. The complex rhythms of CPG are closely related to certain diseases, such as epilepsy. This paper considered the influence of electromagnetic induction on the discharge mode of the half-center oscillators. First, we analyzed the response of individual firing neuron rhythms to electromagnetic induction when the slow-variable parameters vary. We also discussed the changes in the dynamic bifurcation structure when the intensity of electromagnetic induction varies. Furthermore, we determined the effects of mutually inhibitory and self-inhibitory synaptic parameters on the firing rhythm of the half-center oscillators. The different responses induced by electromagnetic induction interventions, showed that mutually inhibitory synapses modulate the firing rhythm weakly and self-inhibition synapses have a significant impact on firing rhythm. Finally, with the change of synaptic parameter values, the combined effects of autapse and mutually inhibitory synapses on the discharge rhythm of half-center oscillators were analyzed in symmetric and asymmetric autapse modes. It was found that the synchronous state of the half-center oscillators had a more robust electromagnetic induction response than the asynchronous state.
Citation: Feibiao Zhan, Jian Song. Complex rhythm and synchronization of half-center oscillators under electromagnetic induction[J]. Electronic Research Archive, 2024, 32(7): 4454-4471. doi: 10.3934/era.2024201
Half-center oscillators are typical small circuits that are crucial for understanding CPG. The complex rhythms of CPG are closely related to certain diseases, such as epilepsy. This paper considered the influence of electromagnetic induction on the discharge mode of the half-center oscillators. First, we analyzed the response of individual firing neuron rhythms to electromagnetic induction when the slow-variable parameters vary. We also discussed the changes in the dynamic bifurcation structure when the intensity of electromagnetic induction varies. Furthermore, we determined the effects of mutually inhibitory and self-inhibitory synaptic parameters on the firing rhythm of the half-center oscillators. The different responses induced by electromagnetic induction interventions, showed that mutually inhibitory synapses modulate the firing rhythm weakly and self-inhibition synapses have a significant impact on firing rhythm. Finally, with the change of synaptic parameter values, the combined effects of autapse and mutually inhibitory synapses on the discharge rhythm of half-center oscillators were analyzed in symmetric and asymmetric autapse modes. It was found that the synchronous state of the half-center oscillators had a more robust electromagnetic induction response than the asynchronous state.

| [1] |
E. Marder, D. Bucher, Central pattern generators and the control of rhythmic movements, Curr. Biol., 11 (2001), R986–R996. https://doi.org/10.1016/S0960-9822(01)00581-4 doi: 10.1016/S0960-9822(01)00581-4

|
| [2] |
E. Marder, R. L. Calabrese, Principles of rhythmic motor pattern generation, Physiol. Rev., 76 (1996), 687–717. https://doi.org/10.1152/physrev.1996.76.3.687 doi: 10.1152/physrev.1996.76.3.687
|
| [3] |
D. N. Masaev, A. A. Suleimanova, N. V. Prudnikov, M. V. Serenko, A. V. Emelyanov, V. A. Demin, et al., Memristive circuit-based model of central pattern generator to reproduce spinal neuronal activity in walking pattern, Front. Neurosci., 17 (2023), 1124950. https://doi.org/10.3389/fnins.2023.1124950 doi: 10.3389/fnins.2023.1124950
|
| [4] |
D. Alaçam, A. Shilnikov, Making a swim central pattern generator out of latent parabolic bursters, Int. J. Bifurcation Chaos, 25 (2015), 1540003. https://doi.org/10.1142/S0218127415400039 doi: 10.1142/S0218127415400039
|
| [5] |
E. Marder, S. Kedia, E. O. Morozova, New insights from small rhythmic circuits, Curr. Opin. Neurobiol., 76 (2022), 102610. https://doi.org/10.1016/j.conb.2022.102610 doi: 10.1016/j.conb.2022.102610
|
| [6] |
E. Marder, Neuromodulation of neuronal circuits: back to the future, Neuron, 76 (2012), 1–11. https://doi.org/10.1016/j.neuron.2012.09.010 doi: 10.1016/j.neuron.2012.09.010
|
| [7] |
T. Nowotny, M. I. Rabinovich, Dynamical origin of independent spiking and bursting activity in neural microcircuits, Phys. Rev. Lett., 98 (2007), 128106. https://doi.org/10.1103/PhysRevLett.98.128106 doi: 10.1103/PhysRevLett.98.128106
|
| [8] |
M. Lodi, A. L. Shilnikov, M. Storace, Design principles for central pattern generators with preset rhythms, IEEE Trans. Neural Networks Learn. Syst., 31 (2019), 3658–3669. https://doi.org/10.1109/TNNLS.2019.2945637 doi: 10.1109/TNNLS.2019.2945637
|
| [9] |
J. T. C. Schwabedal, A. B. Neiman, A. L. Shilnikov, Robust design of polyrhythmic neural circuits, Phys. Rev. E, 90 (2014), 022715. https://doi.org/10.1103/PhysRevE.90.022715 doi: 10.1103/PhysRevE.90.022715
|
| [10] |
J. Collens, K. Pusuluri, A. Kelley, D. Knapper, T. Xing, S. Basodi, et al., Dynamics and bifurcations in multistable 3-cell neural networks, Chaos, 30 (2020), 072101. https://doi.org/10.1063/5.0011374 doi: 10.1063/5.0011374
|
| [11] |
Q. Lu, J. Tian, Synchronization and stochastic resonance of the small-world neural network based on the CPG, Cognit. Neurodyn., 8 (2014), 217–226. https://doi.org/10.1007/s11571-013-9275-8 doi: 10.1007/s11571-013-9275-8
|
| [12] |
Y. Zang, S. Hong, S. E. De, Firing rate-dependent phase responses of Purkinje cells support transient oscillations, eLife, 9 (2020), e60692. https://doi.org/10.7554/eLife.60692 doi: 10.7554/eLife.60692
|
| [13] |
B. S. Gutkin, G. B. Ermentrout, A. D. Reyes, Phase-response curves give the responses of neurons to transient inputs, J. Neurophysiol., 94 (2005), 1623–1635. https://doi.org/10.1152/jn.00359.2004 doi: 10.1152/jn.00359.2004
|
| [14] |
Y. Zang, E. Marder, Neuronal morphology enhances robustness to perturbations of channel densities, PNAS, 120 (2023), e2219049120. https://doi.org/10.1073/pnas.2219049120 doi: 10.1073/pnas.2219049120
|
| [15] | E. M. Izhikevich, Neural excitability, spiking, and bursting, Int. J. Bifurcation Chaos, 10 (2000), 1171–1266. https://doi.org/10.1142/S0218127400000840 |
| [16] |
B. Lu, X. Jiang, Reduced and bifurcation analysis of intrinsically bursting neuron model, Electron. Res. Arch., 31 (2023), 5928–5945. https://doi.org/10.3934/era.2023301 doi: 10.3934/era.2023301
|
| [17] |
F. Zhan, S. Liu, X. Zhang, J. Wang, B. Lu, Mixed-mode oscillations and bifurcation analysis in a pituitary model, Nonlinear Dyn., 94 (2018), 807–826. https://doi.org/10.1007/s11071-018-4395-7 doi: 10.1007/s11071-018-4395-7
|
| [18] |
H. Zhou, B. Lu, H. Gu, X. Wang, Y. Liu, Complex nonlinear dynamics of bursting of thalamic neurons related to Parkinson's disease, Electron. Res. Arch., 32 (2024), 109–133. https://doi.org/10.3934/era.2024006 doi: 10.3934/era.2024006
|
| [19] |
Z. Song, J. Xu, Codimension-two bursting analysis in the delayed neural system with external stimulations, Nonlinear Dyn., 67 (2012), 309–328. https://doi.org/10.1007/s11071-011-9979-4 doi: 10.1007/s11071-011-9979-4
|
| [20] |
W. B. Kristan, Neuronal decision-making circuits, Curr. Biol., 18 (2008), R928–R932. https://doi.org/10.1016/j.cub.2008.07.081 doi: 10.1016/j.cub.2008.07.081
|
| [21] |
K. L. Briggman, W. B. Kristan, Multifunctional pattern-generating circuits, Annu. Rev. Neurosci., 31 (2008), 271–294. https://doi.org/10.1146/annurev.neuro.31.060407.125552 doi: 10.1146/annurev.neuro.31.060407.125552
|
| [22] |
J. Wojcik, J. Schwabedal, R. Clewley, A. L. Shilnikov, Key bifurcations of bursting polyrhythms in 3-cell central pattern generators, PLoS One, 9 (2014), e92918. https://doi.org/10.1371/journal.pone.0092918 doi: 10.1371/journal.pone.0092918
|
| [23] |
C. A. Tassinari, G. Cantalupo, B. Hoegl, P. Cortelli, L. Tassi, S. Francione, et al., Neuroethological approach to frontolimbic epileptic seizures and parasomnias: the same central pattern generators for the same behaviours, Rev. Neurol., 165 (2009), 762–768. https://doi.org/10.1016/j.neurol.2009.08.002 doi: 10.1016/j.neurol.2009.08.002
|
| [24] |
C. A. Tassinari, E. Gardella, G. Cantalupo, G. Rubboli, Relationship of central pattern generators with parasomnias and sleep-related epileptic seizures, Sleep Med. Clin., 7 (2012), 125–134. https://doi.org/10.1016/j.jsmc.2012.01.003 doi: 10.1016/j.jsmc.2012.01.003
|
| [25] |
F. Zhan, J. Song, S. Liu, The influence of synaptic strength and noise on the robustness of central pattern generator, Electron. Res. Arch., 32 (2024), 686–706. https://doi.org/10.3934/era.2024033 doi: 10.3934/era.2024033
|
| [26] |
V. Baruzzi, M. Lodi, M. Storace, A. Shilnikov, Towards more biologically plausible central-pattern-generator models, Phys. Rev. E, 104 (2021), 064405. https://doi.org/10.1103/PhysRevE.104.064405 doi: 10.1103/PhysRevE.104.064405
|
| [27] | R. L. Calabrese, Half-center oscillators underlying rhythmic movements, in The Handbook of Brain Theory and Neural Networks, (1998), 444–447. |
| [28] |
A. Sakurai, P. S. Katz, The central pattern generator underlying swimming in Dendronotus iris: a simple half-center network oscillator with a twist, J. Neurophysiol., 116 (2016), 1728–1742. https://doi.org/10.1152/jn.00150.2016 doi: 10.1152/jn.00150.2016
|
| [29] |
A. Doloc-Mihu, R. L. Calabrese, A database of computational models of a half-center oscillator for analyzing how neuronal parameters influence network activity, J. Biol. Phys., 37 (2011), 263–283. https://doi.org/10.1007/s10867-011-9215-y doi: 10.1007/s10867-011-9215-y
|
| [30] | A. Doloc-Mihu, R. L. Calabrese, Analysis of family structures reveals robustness or sensitivity of bursting activity to parameter variations in a half-center oscillator (HCO) model, eNeuro, 3 (2016). https://doi.org/10.1523/ENEURO.0015-16.2016 |
| [31] |
Z. Song, J. Xu, Multiple switching and bifurcations of in-phase and anti-phase periodic orbits to chaotic coexistence in a delayed half-center CPG oscillator, Nonlinear Dyn., 111 (2023), 16569–16584. https://doi.org/10.1007/s11071-023-08670-w doi: 10.1007/s11071-023-08670-w
|
| [32] |
Z. Song, J. Xu, Multi-coexistence of routes to chaos in a delayed half-center oscillator (DHCO) system, Nonlinear Dyn., 112 (2024), 1469–1486. https://doi.org/10.1007/s11071-023-09089-z doi: 10.1007/s11071-023-09089-z
|
| [33] |
A. J. White, Sensory feedback expands dynamic complexity and aids in robustness against noise, Biol. Cybern., 116 (2022), 267–269. https://doi.org/10.1007/s00422-021-00917-2 doi: 10.1007/s00422-021-00917-2
|
| [34] |
F. Zhan, S. Liu, Response of electrical activity in an improved neuron model under electromagnetic radiation and noise, Front. Comput. Neurosci., 11 (2017), 107. https://doi.org/10.3389/fncom.2017.00107 doi: 10.3389/fncom.2017.00107
|
| [35] | Z. Wang, Y. Yang, L. Duan, Control effects of electromagnetic induction on epileptic seizures, Nonlinear Dyn., 112 (2024). https://doi.org/10.1007/s11071-024-09373-6 |
| [36] |
A. S. Lele, Y. Fang, J. Ting, A. Raychowdhury, Learning to walk: bio-mimetic hexapod locomotion via reinforcement-based spiking central pattern generation, IEEE J. Emerging Sel. Top. Circuits Syst., 10 (2020), 536–545. https://doi.org/10.1109/JETCAS.2020.3033135 doi: 10.1109/JETCAS.2020.3033135
|
| [37] |
T. Sun, Z. Dai, P. Manoonpong, Distributed-force-feedback-based reflex with online learning for adaptive quadruped motor control, Neural Networks, 142 (2021), 410–427. https://doi.org/10.1016/j.neunet.2021.06.001 doi: 10.1016/j.neunet.2021.06.001
|
| [38] |
B. Muthuswamy, Implementing memristor based chaotic circuits, Int. J. Bifurcation Chaos, 20 (2010), 1335–1350. https://doi.org/10.1142/S0218127410026514 doi: 10.1142/S0218127410026514
|
| [39] |
M. Lv, C. Wang, G. Ren, J. Ma, X. Song, Model of electrical activity in a neuron under magnetic flow effect, Nonlinear Dyn., 85 (2016), 1479–1490. https://doi.org/10.1007/s11071-016-2773-6 doi: 10.1007/s11071-016-2773-6
|
| [40] |
F. Zhan, S. Liu, J. Wang, B. Lu, Bursting patterns and mixed-mode oscillations in reduced Purkinje model, Int. J. Mod. Phys. B, 32 (2018), 1850043. https://doi.org/10.1142/S0217979218500431 doi: 10.1142/S0217979218500431
|
| [41] |
D. Terman, J. E. Rubin, A. C. Yew, C. J. Wilson, Activity patterns in a model for the subthalamopallidal network of the basal ganglia, J. Neurosci., 22 (2002), 2963–2976. https://doi.org/10.1523/JNEUROSCI.22-07-02963.2002 doi: 10.1523/JNEUROSCI.22-07-02963.2002
|
| [42] |
F. Su, J. Wang, S. Niu, H. Li, B. Deng, C. Liu, et al., Nonlinear predictive control for adaptive adjustments of deep brain stimulation parameters in basal ganglia–thalamic network, Neural Networks, 98 (2018), 283–295. https://doi.org/10.1016/j.neunet.2017.12.001 doi: 10.1016/j.neunet.2017.12.001
|
| [43] |
J. Song, S. Liu, H. Lin, Model-based quantitative optimization of deep brain stimulation and prediction of Parkinson's states, Neuroscience, 498 (2022), 105–124. https://doi.org/10.1016/j.neuroscience.2022.05.019 doi: 10.1016/j.neuroscience.2022.05.019
|
| [44] |
J. Song, H. Lin, S. Liu, Basal ganglia network dynamics and function: role of direct, indirect and hyper-direct pathways in action selection, Network: Comput. Neural Syst., 34 (2023), 84–121. https://doi.org/10.1080/0954898X.2023.2173816 doi: 10.1080/0954898X.2023.2173816
|
| [45] |
Z. Song, F. Ji, J. Xu, Is there a user-friendly building unit to replicate rhythmic patterns of CPG systems? Synchrony transition and application of the delayed bursting-HCO model, Chaos, Solitons Fractals, 182 (2024), 114820. https://doi.org/10.1016/j.chaos.2024.114820 doi: 10.1016/j.chaos.2024.114820
|









Figures(10)
Feibiao Zhan, Jian Song. Complex rhythm and synchronization of half-center oscillators under electromagnetic induction[J]. Electronic Research Archive, 2024, 32(7): 4454-4471. doi: 10.3934/era.2024201







 DownLoad:
DownLoad: