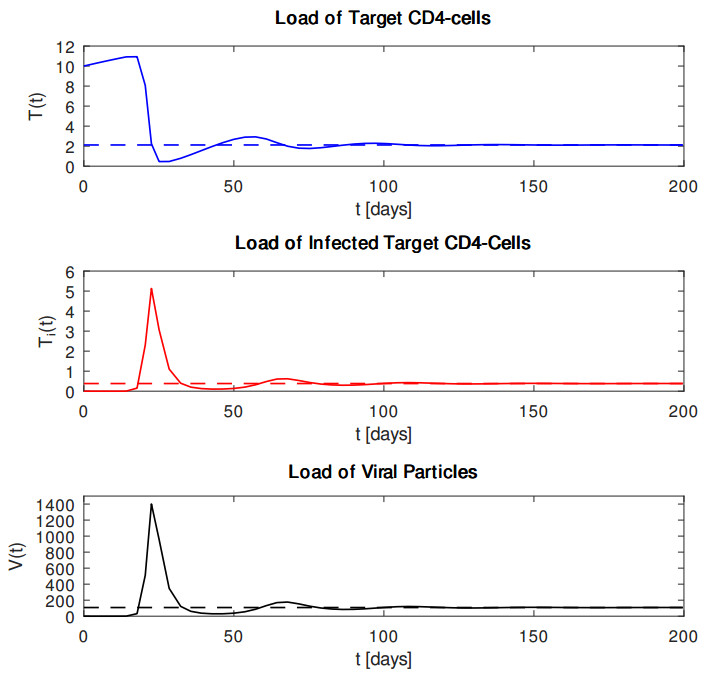
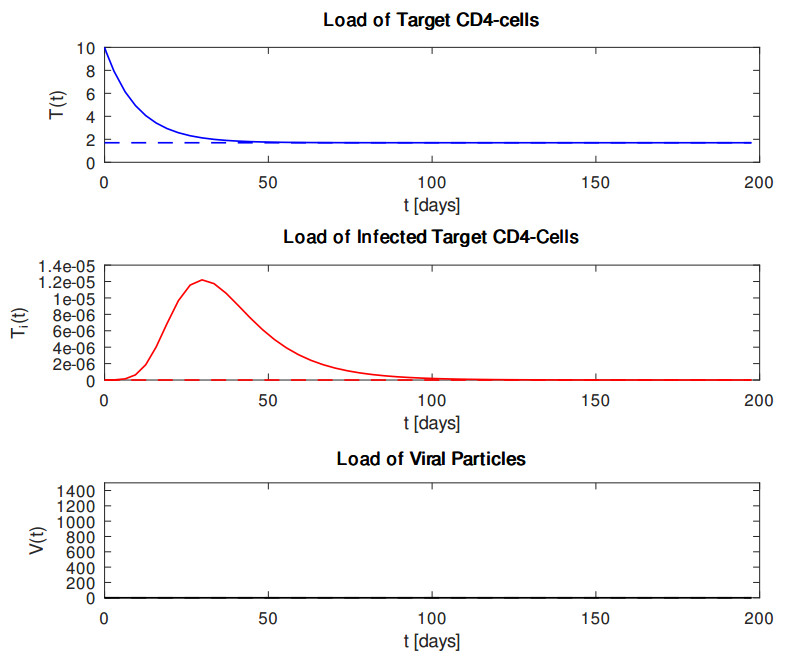
In this article, we reconsider the classical target cell limited dynamical within-host HIV model, solely taking into account the interaction between $ {\rm{CD}}4^{+} $ T cells and virus particles. First, we summarize some analytical results regarding the corresponding dynamical system. For that purpose, we proved some analytical results regarding the system of differential equations as our first main contribution. Specifically, we showed non-negativity and boundedness of solutions, global existence in time and global uniqueness in time and examined stability properties of two possible equilibria. In particular, we demonstrated that the virus-free equilibrium and the plateau-phase equilibrium are locally asymptotically stable using the Routh–Hurwitz criterion under appropriate conditions. As our second main contribution, we underline our theoretical findings through some numerical experiments with standard Runge–Kutta time stepping schemes. We conclude this work with a summary of our main results and a suggestion of an extension for more complex dynamical systems with regard to HIV-infection.
Citation: Benjamin Wacker. Revisiting the classical target cell limited dynamical within-host HIV model - Basic mathematical properties and stability analysis[J]. Mathematical Biosciences and Engineering, 2024, 21(12): 7805-7829. doi: 10.3934/mbe.2024343
In this article, we reconsider the classical target cell limited dynamical within-host HIV model, solely taking into account the interaction between $ {\rm{CD}}4^{+} $ T cells and virus particles. First, we summarize some analytical results regarding the corresponding dynamical system. For that purpose, we proved some analytical results regarding the system of differential equations as our first main contribution. Specifically, we showed non-negativity and boundedness of solutions, global existence in time and global uniqueness in time and examined stability properties of two possible equilibria. In particular, we demonstrated that the virus-free equilibrium and the plateau-phase equilibrium are locally asymptotically stable using the Routh–Hurwitz criterion under appropriate conditions. As our second main contribution, we underline our theoretical findings through some numerical experiments with standard Runge–Kutta time stepping schemes. We conclude this work with a summary of our main results and a suggestion of an extension for more complex dynamical systems with regard to HIV-infection.

| [1] |
A. Mody, A. H. Sohn, C. Iwuji, R. K. J. Tan, F. Venter, E. H. Geng, HIV epidemiology, prevention, treatment, and implementation strategies for public health, Lancet, 402 (2024), 471–492. https://doi.org/10.1016/S0140-6736(23)01381-8 doi: 10.1016/S0140-6736(23)01381-8

|
| [2] | UNAids, Fact Sheet 2024 - Global HIV Statistics, 2024. Available from: https://www.unaids.org/sites/default/files/media_asset/UNAIDS_FactSheet_en.pdf. |
| [3] |
P. Essunger, A. S. Perelson, Modeling HIV infection of $ {\rm{CD}}4^{+}$ T-cell subpopulations, J. Theor. Biol., 170 (1994), 367–391. https://doi.org/10.1006/jtbi.1994.1199 doi: 10.1006/jtbi.1994.1199
|
| [4] |
D. D. Ho, A. U. Neumann, A. S. Perelson, W. Chen, J. M. Leonard, M. Markowitz, Rapid turnover of plasma virions and CD4 lymphocytes in HIV-1 infection, Nature, 373 (1995), 123–126. https://doi.org/10.1038/373123a0 doi: 10.1038/373123a0
|
| [5] | D. Kirschner, Using mathematics to understand HIV immune dynamics, Not. AMS, 43 (1996), 191–202. |
| [6] |
M. A. Nowak, C. R. M. Bangham, Population dynamics of immune responses to persistent viruses, Science, 272 (1996), 74–79. https://doi.org/10.1126/science.272.5258.74 doi: 10.1126/science.272.5258.74
|
| [7] |
S. Bonhoeffer, R. M. May, G. M. Shaw, M. A. Nowak, Virus dynamics and drug therapy, Proc. Natl. Acad. Sci., 94 (1997), 6971–6976. https://doi.org/10.1073/pnas.94.13.6971 doi: 10.1073/pnas.94.13.6971
|
| [8] |
D. Finzi, R. F. Siliciano, Viral dynamics in HIV-1-infection, Cell, 93 (1997), 665–671. https://doi.org/10.1016/s0092-8674(00)81427-0 doi: 10.1016/s0092-8674(00)81427-0
|
| [9] |
R. J. De Boer, A. S. Perelson, Target cell limited and immune control models of HIV infection: A comparison, J. Theor. Biol., 190 (1998), 201–214. https://doi.org/10.1006/jtbi.1997.0548 doi: 10.1006/jtbi.1997.0548
|
| [10] |
A. S. Perelson, P. W. Nelson, Mathematical analysis of HIV-1-dynamics in vivo, SIAM Rev., 41 (1999), 3–44. https://doi.org/10.1137/S0036144598335107 doi: 10.1137/S0036144598335107
|
| [11] |
M. A. Stafford, L. Corey, Y. Cao, E. S. Daar, D. D. Ho, A. S. Perelson, Modeling plasma virus concentration during primary HIV infection, J. Theor. Biol., 203 (1999), 285–301. https://doi.org/10.1006/jtbi.2000.1076 doi: 10.1006/jtbi.2000.1076
|
| [12] |
A. S. Perelson, Modelling viral and immune system dynamics, Nat. Rev. Immunol., 2 (1999), 28–36. https://doi.org/10.1038/nri700 doi: 10.1038/nri700
|
| [13] |
A. Korobeinikov, A Lyapunov function and global properties for SIR and SEIR epidemiological Models with Nonlinear Incidence, Math. Biosci. Eng., 1 (2004), 57–60. https://doi.org/10.3934/mbe.2004.1.57 doi: 10.3934/mbe.2004.1.57
|
| [14] |
A. Korobeinikov, Global properties of basic virus dynamics models, Bull. Math. Biol., 66 (2004), 879–883. https://doi.org/10.1016/j.bulm.2004.02.001 doi: 10.1016/j.bulm.2004.02.001
|
| [15] |
L. Wang, M. Y. Li, Mathematical analysis of the global dynamics of a model for HIV infection of $ {\rm{CD4}}^{+}$ T cells, Math. Biosci., 200 (2006), 44–57. https://doi.org/10.1016/j.mbs.2005.12.026 doi: 10.1016/j.mbs.2005.12.026
|
| [16] |
R. M. Ribeiro, Dynamics of $ {\rm{CD4}}^{+}$ T cells in HIV-1 infection, Immunol. Cell Biol., 85 (2006), 287–294. https://doi.org/10.1038/sj.icb.7100056 doi: 10.1038/sj.icb.7100056
|
| [17] |
S. Alizon, C. Magnus, Modelling the course of an HIV infection: Insights from ecology and evolution, Viruses, 4 (2012), 1984–2013. https://doi.org/10.3390/v4101984 doi: 10.3390/v4101984
|
| [18] |
A. S. Perelson, R. M. Ribeiro, Modeling the within-host dynamics of HIV infection, BMC Biol., 11 (2013), 96. https://doi.org/10.1186/1741-7007-11-96 doi: 10.1186/1741-7007-11-96
|
| [19] |
D. Kirschner, G. Webb, Immunotherapy of HIV-1 infection, J. Biol. Syst., 6 (1998), 71–83. https://doi.org/10.1142/S0218339098000091 doi: 10.1142/S0218339098000091
|
| [20] |
A. Mojaver, H. Kheiri, Mathematical analysis of a class of HIV infection models of CD$4^{+}$ T-cells with combined antiretroviral therapy, Appl. Math. Comput., 259 (2015), 258–270. https://doi.org/10.1016/j.amc.2015.02.064 doi: 10.1016/j.amc.2015.02.064
|
| [21] |
H. F. Huo, R. Chen, X. Y. Wang, Modelling and stability of HIV/AIDS epidemic model with treatment, Appl. Math. Modell., 40 (2016), 6550–6559. https://doi.org/10.1016/j.apm.2016.01.054 doi: 10.1016/j.apm.2016.01.054
|
| [22] |
A. N. Timsina, Y. R. Liyanage, M. Martcheva, N. Tuncer, A novel within-host modelof HIV and nutrition, Math. Biosci. Eng., 21 (2024), 5577–5603. https://doi.org/10.3934/mbe.2024246 doi: 10.3934/mbe.2024246
|
| [23] |
S. Xu, Saturated lysing efficiency of $ {\rm{CD8}}^{+}$ cells induced monostable, bistable and oscillatory HIV kinetics, Math. Biosci. Eng., 21 (2024), 7373–7393. https://doi.org/10.3934/mbe.2024324 doi: 10.3934/mbe.2024324
|
| [24] |
A. L. Jenner, M. Smalley, D. Goldman, W. F. Goins, C. S. Gobbs, R. B. Puchaslki, et al., Agent-based computational modeling of glioblastoma predicts that stromal density is central to oncolytic virus efficacy, iScience, 25 (2022), 104395. https://doi.org/10.1016/j.isci.2022.104395 doi: 10.1016/j.isci.2022.104395
|
| [25] |
A. Surendran, A. L. Jenner, E. Karimi, B. Fiset, D. F. Quail, L. A. Walsh, et al., Agent-based modelling reveals the role of the tumor Microenvironment on the short-term success of combination temozolomide/immune checkpoint blockade to treat glioblastoma, J. Pharmacol. Exp. Ther., 387 (2023), 66–77. https://doi.org/10.1124/jpet.122.001571 doi: 10.1124/jpet.122.001571
|
| [26] |
B. Wacker, J. C. Schlüter, An age- and sex-structured SIR model: Theory and an explicit-implicit numerical solution algorithm, Math. Biosci. Eng., 17 (2020), 5752–5801. https://doi.org/10.3934/mbe.2020309 doi: 10.3934/mbe.2020309
|
| [27] |
B. Wacker, J. C. Schlüter, Time-continuous and time-discrete SIR models revisited: theory and applications, Adv. Differ. Equations, 2020 (2020), 556. 10.1186/s13662-020-02995-1 doi: 10.1186/s13662-020-02995-1
|
| [28] |
B. Wacker, J. C. Schlüter, Qualitative analysis of two systems of nonlinear first-order ordinary differential equations for biological systems, Math. Methods Appl. Sci., 45 (2022), 4597–4624. https://doi.org/10.1002/mma.8056 doi: 10.1002/mma.8056
|
| [29] |
B. Wacker, J. C. Schlüter, A non-standard finite-difference-method for a non-autonomous epidemiological model: analysis, parameter identification and applications, Math. Biosci. Eng., 20 (2023), 12923–12954. https://doi.org/10.3934/mbe.2023577 doi: 10.3934/mbe.2023577
|
| [30] |
B. Wacker, Framework for solving dynamics of $ {\rm{Ca}}^{2+}$ ion concentrations in liver cells numerically: Analysis of a non-negativity-preserving non-standard finite-difference-method, Math. Methods Appl. Sci., 46 (2023), 16625–16643. https://doi.org/10.1002/mma.9464 doi: 10.1002/mma.9464
|
| [31] |
O. Diekmann, J. A. P. Heesterbeek, J. A. J. Metz, On the definition and the computation of the basic reproduction number $R_{0}$ in models for infectious diseases in heterogeneous populations, J. Math. Biol., 28 (1990), 365–382. https://doi.org/10.1007/BF00178324 doi: 10.1007/BF00178324
|
| [32] |
P. van den Driessche, J. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180 (2002), 29–48. https://doi.org/10.1016/S0025-5564(02)00108-6 doi: 10.1016/S0025-5564(02)00108-6
|
| [33] |
I. M. Elbaz, H. El-Metwally, M. A. Sohaly, Viral kinetics, stability and sensitivity analysis of the within-host COVID-19 model, Sci. Rep., 13 (2023), 11675. https://doi.org/10.1038/s41598-023-38705-6 doi: 10.1038/s41598-023-38705-6
|
| [34] | H. K. Khalil, Nonlinear Systems, Prentice-Hall, Upper Saddle River, 2001. |
| [35] | D. G. Schaeffer, J. W. Cain, Ordinary Differential Equations: Basics and Beyond, Springer-Verlag, New York, 2016. https://doi.org/10.1007/978-1-4939-6389-8 |
| [36] |
B. Wacker, J. C. Schlüter, A cubic nonlinear population growth modelfor single species: theory, an explicit-implicit solution algorithm and applications, Adv. Differ. Equations, 2021 (2021), 236. https://doi.org/10.1186/s13662-021-03399-5 doi: 10.1186/s13662-021-03399-5
|
| [37] | M. Reed, B. Simon, Functional Analysis, Academic Press, San Diego, 1980. |
| [38] |
A. Hurwitz, Über die Bedingungen, unter welchen eine Gleichung nur Wurzeln mit negativen reellen Theilen besitzt, Math. Ann., 46 (1895), 273–284. https://doi.org/10.1007/BF01446812 doi: 10.1007/BF01446812
|
| [39] | J. W. Eaton, D. Bateman, S. Hauberg, R. Wehbring, GNU Octave Version 6.1.0 Manual: A High-level Interactive Language for Numerical Computations, 2020. Available from: https://www.gnu.org/software/octave/doc/v6.1.0/. |
| [40] |
J. R. Dormand, P. J. Prince, A family of embedded Runge-Kutta formulae, J. Comput. Appl. Math., 6 (1980), 19–26. https://doi.org/10.1016/0771-050X(80)90013-3 doi: 10.1016/0771-050X(80)90013-3
|
| [41] |
L. F. Shampine, M. W. Reichert, The MATLAB ODE suite, SIAM J. Sci. Comput., 18 (1997), 1–22. https://doi.org/10.1137/S1064827594276424 doi: 10.1137/S1064827594276424
|
| [42] |
T. M. Hoang, High-order nonstandard finite difference methods preserving dynamical properties of one-dimensional dynamical systems, Numerical Algorithms, 2024 (2024), 1–31. https://doi.org/10.1007/s11075-024-01792-1 doi: 10.1007/s11075-024-01792-1
|
| [43] | R. E. Mickens, Nonstandard Finite Difference Models of Differential Equations, World Scientific, 1993. https://doi.org/10.1142/2081 |
 mbe-21-12-343-Supplementary.pdf mbe-21-12-343-Supplementary.pdf |
 |
Figures(2) / Tables(3)

Benjamin Wacker. Revisiting the classical target cell limited dynamical within-host HIV model - Basic mathematical properties and stability analysis[J]. Mathematical Biosciences and Engineering, 2024, 21(12): 7805-7829. doi: 10.3934/mbe.2024343







 DownLoad:
DownLoad: