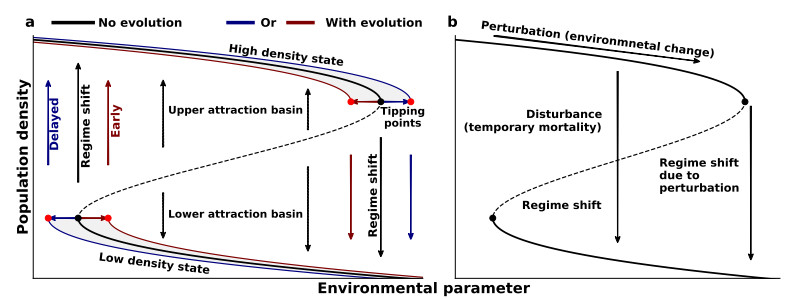
Environmental changes are a growing concern, as they exert pressures on ecosystems. In some cases, such changes lead to shifts in ecosystem structure. However, species can adapt to changes through evolution, and it is unclear how evolution interacts with regime shifts, which restricts ecosystem management strategies. Here, we used a model of prey population with evolution and intra-specific trait diversity, and simulated regime shifts through changes in predation pressure. We then explored interactions between evolution, diversity, and shifts in population density. Evolution induced delayed or early regime shifts, and altered the recovery of populations. Such changes depended on the relative speed of evolution and change of predation pressure, as well as on the initial state of the population. Evolution also influenced population resilience, which was important when considering strong environmental variability. For instance, storms can spontaneously increase mortality and induce shifts. Furthermore, environmental variability induced even higher mortality if the phenotypic diversity of populations is large. Some phenotypes were more vulnerable to environmental changes, and such increases in mortality favor shifts to decreases in density. Thus, population management needs to consider diversity, evolution, and environmental change altogether to better anticipate regime shifts on eco-evolutionary time scales. Here, evolution and diversity showed complex interactions with population shift dynamics. Investigating the influence of higher diversity levels, such as diversity at a community level, should be another step towards anticipating changes in ecosystems and communities.
Citation: Thomas Imbert, Jean-Christophe Poggiale, Mathias Gauduchon. Intra-specific diversity and adaptation modify regime shifts dynamics under environmental change[J]. Mathematical Biosciences and Engineering, 2024, 21(12): 7783-7804. doi: 10.3934/mbe.2024342
Environmental changes are a growing concern, as they exert pressures on ecosystems. In some cases, such changes lead to shifts in ecosystem structure. However, species can adapt to changes through evolution, and it is unclear how evolution interacts with regime shifts, which restricts ecosystem management strategies. Here, we used a model of prey population with evolution and intra-specific trait diversity, and simulated regime shifts through changes in predation pressure. We then explored interactions between evolution, diversity, and shifts in population density. Evolution induced delayed or early regime shifts, and altered the recovery of populations. Such changes depended on the relative speed of evolution and change of predation pressure, as well as on the initial state of the population. Evolution also influenced population resilience, which was important when considering strong environmental variability. For instance, storms can spontaneously increase mortality and induce shifts. Furthermore, environmental variability induced even higher mortality if the phenotypic diversity of populations is large. Some phenotypes were more vulnerable to environmental changes, and such increases in mortality favor shifts to decreases in density. Thus, population management needs to consider diversity, evolution, and environmental change altogether to better anticipate regime shifts on eco-evolutionary time scales. Here, evolution and diversity showed complex interactions with population shift dynamics. Investigating the influence of higher diversity levels, such as diversity at a community level, should be another step towards anticipating changes in ecosystems and communities.

| [1] |
V. Dakos, S. R. Carpenter, W. A. Brock, A. M. Ellison, V. Guttal, A. R. Ives, et al., Methods for detecting early warnings of critical transitions in time series illustrated using simulated ecological data, PLOS One, 7 (2012), e41010. https://doi.org/10.1371/journal.pone.0041010 doi: 10.1371/journal.pone.0041010

|
| [2] |
J. C. Blackwood, A. Hastings, P. J. Mumby, The effect of fishing on hysteresis in Caribbean coral reefs, Theor. Ecol., 5 (2012), 105–114. https://doi.org/10.1007/s12080-010-0102-0 doi: 10.1007/s12080-010-0102-0
|
| [3] |
B. E. Beisner, D. T. Haydon, K. Cuddington, Alternative stable states in ecology, Front. Ecol. Environ., 1 (2012), 376–382. https://doi.org/10.1890/1540-9295(2003)001[0376:ASSIE]2.0.CO;2 doi: 10.1890/1540-9295(2003)001[0376:ASSIE]2.0.CO;2
|
| [4] |
T. Yoshida, L. E. Jones, S. P. Ellner, G. F. Fussmann, N. G. Hairston, Rapid evolution drives ecological dynamics in a predator–prey system, Nature, 424 (2003), 303–306. https://doi.org/10.1038/nature01767 doi: 10.1038/nature01767
|
| [5] |
R. Dawkins, J. R. Krebs, Arms races between and within species, Proc. R. Soc. Lond. B., 205 (1979), 485–511. https://doi.org/10.1098/rspb.1979.0081 doi: 10.1098/rspb.1979.0081
|
| [6] |
O. Cotto, L. Sandell, L. M. Chevin, O. Ronce, Maladaptive shifts in life history in a changing environment, Am. Nat., 194 (2019), 558–573. https://doi.org/10.1086/702716 doi: 10.1086/702716
|
| [7] |
V. Dakos, B. Matthews, A. P. Hendry, J. Levine, N. Loeuille, J. Norberg, et al., Ecosystem tipping points in an evolving world, Nat. Ecol. Evol., 3 (2019), 355–362. https://doi.org/10.1038/s41559-019-0797-2 doi: 10.1038/s41559-019-0797-2
|
| [8] |
F. Dercole, R. Ferriere, S. Rinaldi, Ecological bistability and evolutionary reversals under asymmetrical competition, Evolution, 56 (2002), 1081–1090. https://doi.org/10.1111/j.0014-3820.2002.tb01422.x doi: 10.1111/j.0014-3820.2002.tb01422.x
|
| [9] |
M. G. Bulmer, The effect of selection on genetic variability, Am. Nat., 105 (2008), 201–211. https://doi.org/10.1086/282718 doi: 10.1086/282718
|
| [10] |
A. Ehlers, B. Worm, T. B. H. Reusch, Importance of genetic diversity in eelgrass Zostera marina for its resilience to global warming, Mar. Ecol. Prog. Ser., 365 (2008), 1–7. https://doi.org/10.3354/meps07369 doi: 10.3354/meps07369
|
| [11] |
C. M. Sgrò, A. J. Lowe, A. A. Hoffmann, Building evolutionary resilience for conserving biodiversity under climate change, Evol. Appl., 4 (2011), 326–337. https://doi.org/10.1111/j.1752-4571.2010.00157.x doi: 10.1111/j.1752-4571.2010.00157.x
|
| [12] | M. C. Donaldson-Matasci, M. Lachmann, C. T. Bergstrom, Phenotypic diversity as an adaptation to environmental uncertainty, Evol. Ecol. Res., 10 (2008), 493–515. https://www.evolutionary-ecology.com/abstracts/v10/2266.html |
| [13] |
A. N. Ardichvili, N. Loeuille, V. Dakos, Evolutionary emergence of alternative stable states in shallow lakes, Ecol. Lett., 26 (2022), 692–705. https://doi.org/10.1111/ele.14180 doi: 10.1111/ele.14180
|
| [14] |
J. Garnier, O. Cotto, E. Bouin, T. Bourgeron, T. Lepoutre, O. Ronce, et al., Adaptation of a quantitative trait to a changing environment: New analytical insights on the asexual and infinitesimal sexual models, Theor. Popul. Biol., 152 (2023), 1–22. https://doi.org/10.1016/j.tpb.2023.04.002 doi: 10.1016/j.tpb.2023.04.002
|
| [15] |
S. A. H. Geritz, J. A. J. Metz, E. Kisdi, G. Meszéna, Dynamics of adaptation and evolutionary branching, Phys. Rev. Lett., 78 (1997), 2024–2027. https://doi.org/10.1103/PhysRevLett.78.2024 doi: 10.1103/PhysRevLett.78.2024
|
| [16] |
C. S. Holling, Resilience and stability of ecological systems, Annu. Rev. Ecol. Syst., 4 (1973), 1–23. https://doi.org/10.1146/annurev.es.04.110173.000245 doi: 10.1146/annurev.es.04.110173.000245
|
| [17] |
M. Scheffer, S. Carpenter, J. A. Foley, C. Folke, B. Walker, Catastrophic shifts in ecosystems, Nature, 413 (2001), 591–596. https://doi.org/10.1038/35098000 doi: 10.1038/35098000
|
| [18] |
G. Booy, R. J. J. Hendriks, M. J. M. Smulders, J. M. Van Groenendael, B. Vosman, Genetic diversity and the survival of populations, Plant Biol., 2 (2000), 379–395. https://doi.org/10.1055/s-2000-5958 doi: 10.1055/s-2000-5958
|
| [19] |
D. Ludwig, D. D. Jones, C. S. Holling, Qualitative analysis of insect outbreak systems: The spruce budworm and forest, J. Anim. Ecol., 47 (1978), 315–332. https://doi.org/10.2307/3939 doi: 10.2307/3939
|
| [20] | É. Kisdi, Evolutionary branching under asymmetric competition, J. Theor. Biol., 197 (1999), 149–162. https://linkinghub.elsevier.com/retrieve/pii/S0022519398908645 |
| [21] |
L. M. Guzman, D. S. Srivastava, Prey body mass and richness underlie the persistence of a top predator, Proc. R. Soc. B, 286 (2019), 20190622. https://doi.org/10.1098/rspb.2019.0622 doi: 10.1098/rspb.2019.0622
|
| [22] |
P. G. Milonas, D. C. Kontodimas, A. F. Martinou, A predator's functional response: Influence of prey species and size, Biol. Control, 59 (2011), 141–146. https://doi.org/10.1016/j.biocontrol.2011.06.016 doi: 10.1016/j.biocontrol.2011.06.016
|
| [23] |
D. J. Rankin, A. López‐Sepulcre, Can adaptation lead to extinction?, Oikos, 111 (2005), 616–619. https://doi.org/10.1111/j.1600-0706.2005.14541.x doi: 10.1111/j.1600-0706.2005.14541.x
|
| [24] |
B. Perthame, M. Gauduchon, Survival thresholds and mortality rates in adaptive dynamics: Conciliating deterministic and stochastic simulations, Math. Med. Biol., 27 (2010), 195–210. https://doi.org/10.1093/imammb/dqp018 doi: 10.1093/imammb/dqp018
|
| [25] | M. A. McPeek, The ecological dynamics of natural selection: Traits and the coevolution of community structure, Am. Nat., 189 (2017), E91–E117. http://orcid.org/0000-0001-7794-9466 |
| [26] |
J. M. Chase, P. A. Abrams, J. P. Grover, S. Diehl, P. Chesson, R. D. Holt, et al., The interaction between predation and competition: A review and synthesis, Ecol. Lett., 5 (2002), 302–315. https://doi.org/10.1046/j.1461-0248.2002.00315.x doi: 10.1046/j.1461-0248.2002.00315.x
|
| [27] |
J. Gurevitch, J. A. Morrison, L. V. Hedges, The interaction between competition and predation: A meta‐analysis of field experiments, Am. Nat., 155 (2000), 435–453. https://doi.org/10.1086/303337 doi: 10.1086/303337
|
| [28] |
J. Roughgarden, M. Feldman, Species packing and predation pressure, Ecology, 56 (1975), 489–492. https://doi.org/10.2307/1934982 doi: 10.2307/1934982
|
| [29] |
M. Haimovici, L. M. Cavole, J. M. Cope, L. G. Cardoso, Long-term changes in population dynamics and life history contribute to explain the resilience of a stock of Micropogonias furnieri (Sciaenidae, Teleostei) in the SW Atlantic, Fish. Res., 237 (2021), 105878. https://doi.org/10.1016/j.fishres.2021.105878 doi: 10.1016/j.fishres.2021.105878
|
| [30] |
P. C. Chaparro-Pedraza, Fast environmental change and eco-evolutionary feedbacks can drive regime shifts in ecosystems before tipping points are crossed, Proc. R. Soc. B, 288 (2021), 20211192. https://doi.org/10.1098/rspb.2021.1192 doi: 10.1098/rspb.2021.1192
|
| [31] |
H. Kokko, A. López-Sepulcre, The ecogenetic link between demography and evolution: Can we bridge the gap between theory and data?, Ecol. Lett., 10 (2007), 773–782. https://doi.org/10.1111/j.1461-0248.2007.01086.x doi: 10.1111/j.1461-0248.2007.01086.x
|
| [32] |
J. J. Bull, J. Millstein, J. Orcutt, H. A. Wichman, Evolutionary feedback mediated through population density, illustrated with viruses in chemostats, Am. Nat., 167 (2006), E39–E51. https://doi.org/10.1086/499374 doi: 10.1086/499374
|
| [33] |
L. Přibylová, Regime shifts caused by adaptive dynamics in prey–predator models and their relationship with intraspecific competition, Ecol. Complex., 36 (2018), 48–56. https://doi.org/10.1016/j.ecocom.2018.06.003 doi: 10.1016/j.ecocom.2018.06.003
|
| [34] |
M. H. Cortez, Comparing the qualitatively different effects rapidly evolving and rapidly induced defences have on predator–prey interactions, Ecol. Lett., 14 (2011), 202–209. https://doi.org/10.1111/j.1461-0248.2010.01572.x doi: 10.1111/j.1461-0248.2010.01572.x
|
| [35] |
R. Ferriere, S. Legendre, Eco-evolutionary feedbacks, adaptive dynamics and evolutionary rescue theory, Phil. Trans. R. Soc. B, 368 (2013), 20120081. https://doi.org/10.1098/rstb.2012.0081 doi: 10.1098/rstb.2012.0081
|
| [36] |
M. Scheffer, S. M. Carpenter, T. M. Lenton, J. Bascompte, W. Brock, V. Dakos, et al., Anticipating critical transitions, Science, 338 (2012), 344–348. https://doi.org/10.1126/science.1225244 doi: 10.1126/science.1225244
|
| [37] |
K. A. M. Engelhardt, M. W. Lloyd, M. C. Neel, Effects of genetic diversity on conservation and restoration potential at individual, population, and regional scales, Biol. Conserv., 179 (2014), 6–16. https://doi.org/10.1016/j.biocon.2014.08.011 doi: 10.1016/j.biocon.2014.08.011
|
| [38] |
T. H. Oliver, M. S. Heard, N. J. B. Isaac, D. B. Roy, D. Procter, F. Eigenbrod, et al., Biodiversity and resilience of ecosystem functions, Trends Ecol. Evol., 30 (2015), 673–684. https://doi.org/10.1016/j.tree.2015.08.009 doi: 10.1016/j.tree.2015.08.009
|
| [39] |
M. M. Osmond, C. S. Klausmeier, An evolutionary tipping point in a changing environment, Evolution, 71 (2017), 2930–2941. https://doi.org/10.1111/evo.13374 doi: 10.1111/evo.13374
|
| [40] |
H. K. Plaisted, A. B. Novak, S. Weigel, A. S. Klein, F. T. Short, Eelgrass genetic diversity influences resilience to stresses associated with eutrophication, ESCO, 43 (2020), 1425–1438. https://doi.org/10.1007/s12237-019-00669-0 doi: 10.1007/s12237-019-00669-0
|
| [41] |
W. F. Basener, J. C. Sanford, The fundamental theorem of natural selection with mutations, J. Math. Biol., 76 (2018), 1589–1622. https://doi.org/10.1007/s00285-017-1190-x doi: 10.1007/s00285-017-1190-x
|
| [42] |
R. Barrett, D. Schluter, Adaptation from standing genetic variation, Trends Ecol. Evol., 23 (2008), 38–44. https://doi.org/10.1016/j.tree.2007.09.008 doi: 10.1016/j.tree.2007.09.008
|
| [43] |
D. Ortiz-Barrientos, J. Engelstädter, L. H. Rieseberg, Recombination rate evolution and the origin of species, Trends Ecol. Evol., 31 (2016), 226–236. https://doi.org/10.1016/j.tree.2015.12.016 doi: 10.1016/j.tree.2015.12.016
|
| [44] |
M. Loreau, Linking biodiversity and ecosystems: Towards a unifying ecological theory, Phil. Trans. R. Soc. B, 365 (2010), 49–60. https://doi.org/10.1098/rstb.2009.0155 doi: 10.1098/rstb.2009.0155
|
| [45] |
D. M. Post, E. P. Palkovacs, Eco-evolutionary feedbacks in community and ecosystem ecology: Interactions between the ecological theatre and the evolutionary play, Phil. Trans. R. Soc. B, 364 (2009), 1629–1640. https://doi.org/10.1098/rstb.2009.0012 doi: 10.1098/rstb.2009.0012
|
| [46] |
P. C. Chaparro-Pedraza, A. M. De Roos, Ecological changes with minor effect initiate evolution to delayed regime shifts, Nat. Ecol. Evol., 4 (2020), 412–418. https://doi.org/10.1038/s41559-020-1110-0 doi: 10.1038/s41559-020-1110-0
|
| [47] |
T. Lenormand, Gene flow and the limits to natural selection, Trends Ecol. Evol., 17 (2002), 183–189. https://doi.org/10.1016/S0169-5347(02)02497-7 doi: 10.1016/S0169-5347(02)02497-7
|
| [48] |
S. Våge, G. Bratbak, J. Egge, M. Heldal, A. Larsen, S. Norland, et al., Simple models combining competition, defence and resource availability have broad implications in pelagic microbial food webs, Ecol. Lett., 21 (2018), 1440–1452. https://doi.org/10.1111/ele.13122 doi: 10.1111/ele.13122
|
| [49] |
X. Duan, J. Ye, Y. Lu, C. Du, B. Jang, J. Park, Does cooperation among conspecifics facilitate the coexistence of species?, Chaos Solit. Fractals, 186 (2024), 115308. https://doi.org/10.1016/j.chaos.2024.115308 doi: 10.1016/j.chaos.2024.115308
|
| [50] |
S. Patel, M. H. Cortew, S. J. Schreiber, Partitioning the effects of eco-evolutionary feedbacks on community stability, Am. Nat., 191 (2018), 381–394. https://doi.org/10.1086/695834 doi: 10.1086/695834
|
| [51] |
F. Dercole, J. O. Irisson, S. Rinaldi, Bifurcation analysis of a prey-predator coevolution model, SIAM J. Appl. Math., 63 (2003), 1378–1391. http://epubs.siam.org/doi/10.1137/S0036139902411612 doi: 10.1137/S0036139902411612
|
| [52] |
T. Nakazawa, Ontogenetic niche shifts matter in community ecology: A review and future perspectives, Popul. Ecol., 57 (2015), 347–354. https://doi.org/10.1007/s10144-014-0448-z doi: 10.1007/s10144-014-0448-z
|
| [53] |
J. R. King, J. A. McFarlane, A. E. Punt, Shifts in fisheries management: Adapting to regime shifts, Phil. Trans. R. Soc. B, 370 (2015), 20130277. https://doi.org/10.1098/rstb.2013.0277 doi: 10.1098/rstb.2013.0277
|
 mbe-21-12-342-supplementary.pdf mbe-21-12-342-supplementary.pdf |
 |





Figures(6)

Thomas Imbert, Jean-Christophe Poggiale, Mathias Gauduchon. Intra-specific diversity and adaptation modify regime shifts dynamics under environmental change[J]. Mathematical Biosciences and Engineering, 2024, 21(12): 7783-7804. doi: 10.3934/mbe.2024342







 DownLoad:
DownLoad: