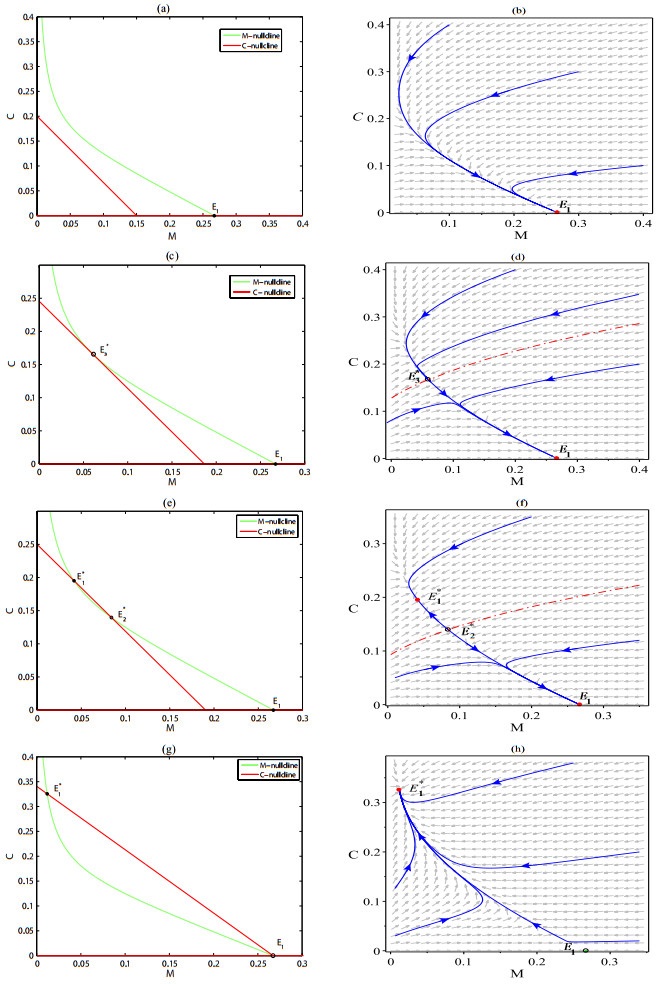
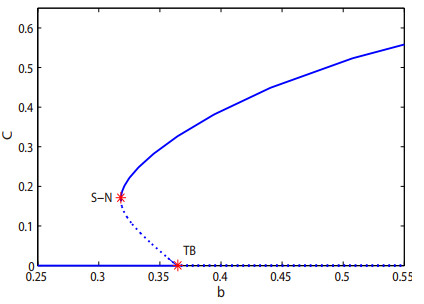
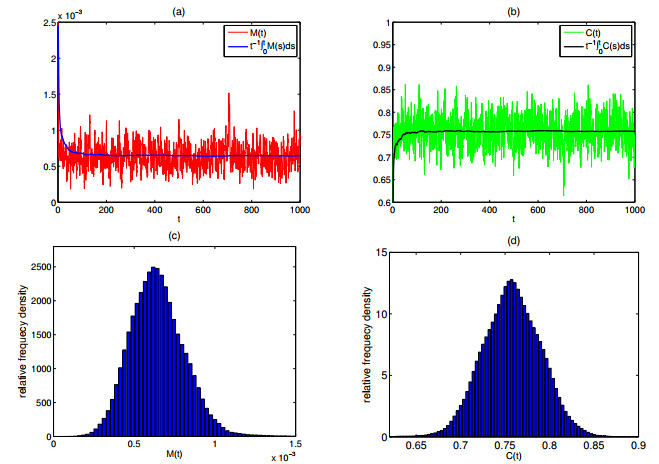
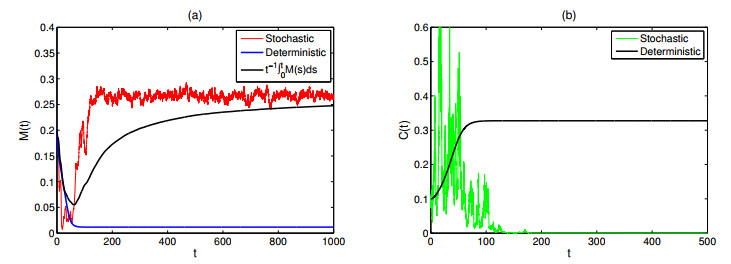
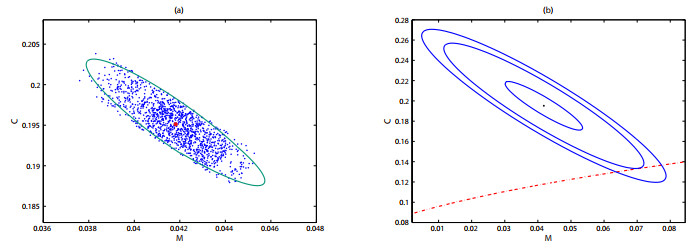
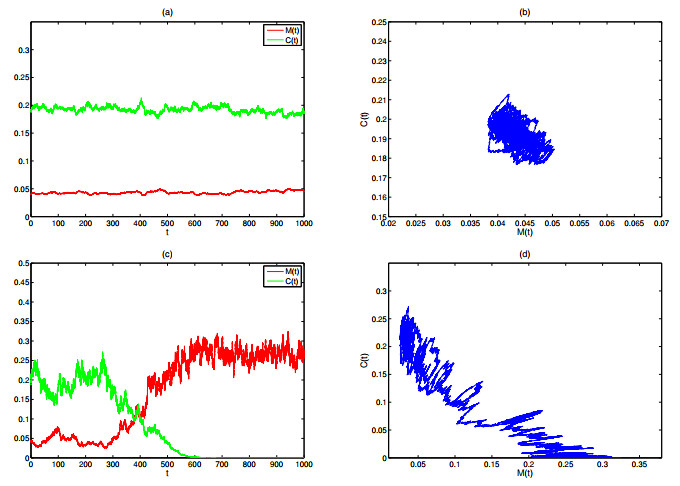
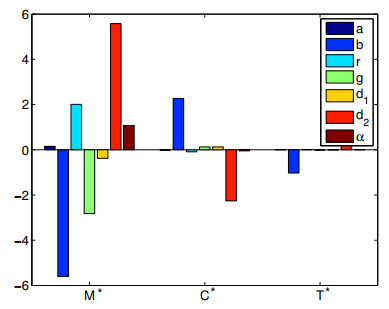
As the most diverse, productive but vulnerable marine habitats in the world, coral reefs are easily affected by the ubiquitous environmental fluctuations, which could change the population dynamics and induce phase shifts of the ecosystem. In this paper, we consider a coral reef benthic system, where macroalgae, corals and algal turfs compete for the available space on a given region of the seabed with grazing intensity and immigrated macroalgae in both deterministic and stochastic environments. For the deterministic system, we analyze the existence and stability of equilibria, as well as the existence of bifurcations. For the stochastic system, sufficient conditions for the existence of the unique ergodic stationary distribution as well as the extinction of corals are obtained, by choosing suitable Lyapunov functions. Moreover, for the scenario that the system exhibits bistability between a macroalgal-coral coexistence equilibrium and a coral-free equilibrium in the absence of environmental fluctuation, we further investigate the irreversible noise-induced transition from macroalgal-coral coexistence to coral extirpation, and numerically estimate the critical values of noise intensity for the occurrence of such transition with the aids of the technique of stochastic sensitivity functions.
Citation: Shengnan Zhao, Sanling Yuan. A coral reef benthic system with grazing intensity and immigrated macroalgae in deterministic and stochastic environments[J]. Mathematical Biosciences and Engineering, 2022, 19(4): 3449-3471. doi: 10.3934/mbe.2022159
As the most diverse, productive but vulnerable marine habitats in the world, coral reefs are easily affected by the ubiquitous environmental fluctuations, which could change the population dynamics and induce phase shifts of the ecosystem. In this paper, we consider a coral reef benthic system, where macroalgae, corals and algal turfs compete for the available space on a given region of the seabed with grazing intensity and immigrated macroalgae in both deterministic and stochastic environments. For the deterministic system, we analyze the existence and stability of equilibria, as well as the existence of bifurcations. For the stochastic system, sufficient conditions for the existence of the unique ergodic stationary distribution as well as the extinction of corals are obtained, by choosing suitable Lyapunov functions. Moreover, for the scenario that the system exhibits bistability between a macroalgal-coral coexistence equilibrium and a coral-free equilibrium in the absence of environmental fluctuation, we further investigate the irreversible noise-induced transition from macroalgal-coral coexistence to coral extirpation, and numerically estimate the critical values of noise intensity for the occurrence of such transition with the aids of the technique of stochastic sensitivity functions.

| [1] | L. Carvajal, Impacts of Climate Change on Human Development, 2007. Available from: https://EconPapers.repec.org/RePEc:hdr:hdocpa:hdocpa-2007-18. |
| [2] | J. L. Munro, The scope of tropical reef fisheries and their management, reef fisheries, in Reef Fisheries, Springer, (1996), 1–14. https://doi.org/10.1007/978-94-015-8779-2_1 |
| [3] | UNEP, Marine and Coastal Ecosystems And Human Well-Being: A Synthesis Report Based On The Findings Of The Millennium Ecosystem Assessment, 2006. Available from: https://www.millenniumassessment.org/documents/Document.799.aspx.pdf. |
| [4] |
N. A. J. Graham, S. K. Wilson, S. Jennings, N. V. C. Polunin, J. P. Bijoux, J. Robinson, Dynamic fragility of oceanic coral reef ecosystems, Proc. Natl. Acad. Sci. USA, 103 (2006), 8425–8429. https://doi.org/10.1073/pnas.0600693103 doi: 10.1073/pnas.0600693103

|
| [5] |
J. C. Blackwood, A. Hastings, P. J. Mumby, The effect of fishing on hysteresis in caribbean coral reefs, Theor. Ecol., 5 (2012), 105–114. https://doi.org/10.1007/s12080-010-0102-0 doi: 10.1007/s12080-010-0102-0
|
| [6] | S. Pal, J. Bhattacharyya, Resilience and dynamics of coral reefs impacted by chemically rich seaweeds and unsustainable fishing, in Mathematical Modelling, Optimization, Analytic and Numerical Solutions, 2020. https://doi.org/10.1007/978-981-15-0928-5_12 |
| [7] |
T. Elmhirst, S. R. Connolly, T. P. Hughes, Connectivity, regime shifts and the resilience of coral reefs, Coral Reefs, 28 (2009), 949–957. https://doi.org/10.1007/s00338-009-0530-8 doi: 10.1007/s00338-009-0530-8
|
| [8] |
P. J. Mumby, N. L. Foster, E. A. G. Fahy, Patch dynamics of coral reef macroalgae under chronic and acute disturbance, Coral Reefs, 24 (2005), 681–692. https://doi.org/10.1007/s00338-005-0058-5 doi: 10.1007/s00338-005-0058-5
|
| [9] |
J. Bhattacharyya, S. Pal, Hysteresis in coral reefs under macroalgal toxicity and overfishing, J. Biol. Phys., 41 (2015), 151–172. https://doi.org/10.1007/s10867-014-9371-y doi: 10.1007/s10867-014-9371-y
|
| [10] |
S. J. Box, P. J. Mumby, The effect of macroalgal competition on the growth and survival of juvenile caribbean corals, Mar. Ecol. Prog. Ser., 342 (2007), 139–149. https://doi.org/10.3354/meps342139 doi: 10.3354/meps342139
|
| [11] |
D. R. Bellwood, T. P. Hughes, C. Folke, M. Nyström, Confronting the coral reef crisis, Nature, 429 (2004), 827–833. https://doi.org/10.1038/nature02691 doi: 10.1038/nature02691
|
| [12] |
J. Jompa, L. J. McCook, Effects of competition and herbivory on interactions between a hard coral and a brown alga, J. Exp. Mar. Biol. Ecol., 271 (2002), 25–39. https://doi.org/10.1016/S0022-0981(02)00040-0 doi: 10.1016/S0022-0981(02)00040-0
|
| [13] |
A. J. Cheal, M. A. MacNeil, E. Cripps, M. J. Emslie, M. Jonker, B. Schaffelke, et al., Coral-macroalgal phase shifts or reef resilience: links with diversity and functional roles of herbivorous fishes on the great barrier reef, Coral Reefs, 29 (2010), 1005–1015. https://doi.org/10.1007/s00338-010-0661-y doi: 10.1007/s00338-010-0661-y
|
| [14] | C. L. Birrell, L. J. Mccook, B. L. Willis G. A. Diaz-Pulido, Effects of benthic algae on the replenishment of corals and the implications for the resilience of coral reefs, in Oceanography and Marine Biology, CRC Press, (2008), 25–63. |
| [15] |
N. Knowlton, Thresholds and multiple stable states in coral reef community dynamics, Am. Zool., 32 (1992), 674–682. https://doi.org/10.1093/icb/32.6.674 doi: 10.1093/icb/32.6.674
|
| [16] |
R. Cropp, J. Norbury, The potential for coral reefs to adapt to a changing climate-an eco-evolutionary modelling perspective, Ecol. Model., 426 (2020), 109038. https://doi.org/10.1016/j.ecolmodel.2020.109038 doi: 10.1016/j.ecolmodel.2020.109038
|
| [17] |
P. J. Mumby, A. Hastings, H. J. Edwards, Thresholds and the resilience of caribbean coral reefs, Nature, 450 (2007), 98–101. https://doi.org/10.1038/nature06252 doi: 10.1038/nature06252
|
| [18] |
J. Yang, S. Yuan, Dynamics of a toxic producing phytoplankton-zooplankton model with three-dimensional patch, Appl. Math. Lett., 118 (2021), 107146. https://doi.org/10.1016/j.aml.2021.107146 doi: 10.1016/j.aml.2021.107146
|
| [19] |
A. Majumder, D. Adak, N. Bairagi, Phytoplankton-zooplankton interaction under environmental stochasticity: survival, extinction and stability, Appl. Math. Model., 89 (2020), 1382–1404. https://doi.org/10.1016/j.apm.2020.06.076 doi: 10.1016/j.apm.2020.06.076
|
| [20] |
H. Wang, M. Liu, Stationary distribution of a stochastic hybrid phytoplankton-zooplankton model with toxin-producing phytoplankton, Appl. Math. Lett., 101 (2020), 106077. https://doi.org/10.1016/j.aml.2019.106077 doi: 10.1016/j.aml.2019.106077
|
| [21] |
D. Jiang, N. Shi, A note on nonautonomous logistic equation with random perturbation, J. Math. Anal. Appl., 303 (2005), 164–172. https://doi.org/10.1016/j.jmaa.2004.08.027 doi: 10.1016/j.jmaa.2004.08.027
|
| [22] |
X. Yu, S. Yuan, T. Zhang, Asymptotic properties of stochastic nutrient-plankton food chain models with nutrient recycling, Nonlinear Anal. Hybri. Syst., 34 (2019), 209–225. https://doi.org/10.1016/j.nahs.2019.06.005 doi: 10.1016/j.nahs.2019.06.005
|
| [23] |
D. Valenti, G. Denaro, B. Spagnolo, S. Mazzola, G. Basilone, F. Conversano, et al., Stochastic models for phytoplankton dynamics in mediterranean sea, Ecol. Complex., 27 (2016), 84–103. https://doi.org/10.1016/j.ecocom.2015.06.001 doi: 10.1016/j.ecocom.2015.06.001
|
| [24] |
D. Song, M. Fan, S. Yan, M. Liu, Dynamics of a nutrient-phytoplankton model with random phytoplankton mortality, J. Theor. Biol., 488 (2019), 110119. https://doi.org/10.1016/j.jtbi.2019.110119 doi: 10.1016/j.jtbi.2019.110119
|
| [25] |
S. Pan, Q. Zhang, A. Meyer-Baese, Stationary distribution of a stochastic vegetation-water system with reaction-diffusion, Appl. Math. Lett., 123 2022, 107589. https://doi.org/10.1016/j.aml.2021.107589 doi: 10.1016/j.aml.2021.107589
|
| [26] |
S. Zhang, S. Yuan, T. Zhang, A predator-prey model with different response functions to juvenile and adult prey in deterministic and stochastic environments, Appl. Math. Comput., 413 (2022), 126598. https://doi.org/10.1016/j.amc.2021.126598 doi: 10.1016/j.amc.2021.126598
|
| [27] |
H. Qi, X. Meng, Threshold behavior of a stochastic predator-prey system with prey refuge and fear effect, Appl. Math. Lett., 113 (2021), 106846. https://doi.org/10.1016/j.aml.2020.106846 doi: 10.1016/j.aml.2020.106846
|
| [28] |
S. Zhang, T. Zhang, S. Yuan, Dynamics of a stochastic predator-prey model with habitat complexity and prey aggregation, Ecol. Complexity, 45 (2021), 100889. https://doi.org/10.1016/j.ecocom.2020.100889 doi: 10.1016/j.ecocom.2020.100889
|
| [29] |
C. Xu, S. Yuan, T. Zhang, Competitive exclusion in a general multi-species chemostat model with stochastic perturbations, Bull. Math. Biol., 83 (2021), 1–17. https://doi.org/10.1007/s11538-020-00843-7 doi: 10.1007/s11538-020-00843-7
|
| [30] |
X. Yu, S. Yuan, Asymptotic properties of a stochastic chemostat model with two distributed delays and nonlinear perturbation, Discrete Cont. Dyn. Syst. B, 25 (2020), 2273–2290. http://dx.doi.org/10.3934/dcdsb.2020014 doi: 10.3934/dcdsb.2020014
|
| [31] | L. Perko, Differential Equations And Dynamical Systems, Springer, 2001. https://doi.org/10.1007/978-1-4613-0003-8 |
| [32] | X. Mao, Stochastic Differential Equations And Applications, Chichester, Horwood, 1997. |
| [33] | R. Khasminskii, Stochastic Stability Of Differential Equations, Springer-Verlag Berlin Heidelberg, 2012. https://doi.org/10.1007/978-3-642-23280-0 |
| [34] |
I. Bashkirtseva, T. Ryazanova, L. Ryashko, Confidence domains in the analysis of noise-induced transition to chaos for Goodwin model of business cycles, Int. J. Bifurcat. Chaos, 24 (2014), 1440020. https://doi.org/10.1142/S0218127414400203 doi: 10.1142/S0218127414400203
|
| [35] |
S. Yuan, D. Wu, G. Lan, H. Wang, Noise-induced transitions in a nonsmooth producer-grazer model with stoichiometric constraints, Bull. Math. Biol., 82 (2020), 55. https://doi.org/10.1007/s11538-020-00733-y doi: 10.1007/s11538-020-00733-y
|
| [36] |
J. Bao, C. Yuan, Stochastic population dynamics driven by l$\acute{\mathrm{e}}$vy noise, J. Math. Anal. Appl., 391 (2012), 363–375. https://doi.org/10.1016/j.jmaa.2012.02.043 doi: 10.1016/j.jmaa.2012.02.043
|
| [37] |
T. Ma, X. Meng, Z. Chang, Dynamics and optimal harvesting control for a stochastic one-predator-two-prey time delay system with jumps, Complexity, 2019 (2019), 1–19. https://doi.org/10.1155/2019/5342031 doi: 10.1155/2019/5342031
|
| [38] |
C. D. Lefevre, D. R. Bellwood, Seasonality and dynamics in coral reef macroalgae: variation in condition and susceptibility to herbivory, Mar. Biol., 157 (2010), 955–965. https://doi.org/10.1007/s00227-009-1376-x doi: 10.1007/s00227-009-1376-x
|
| [39] |
G. Diaz-Pulido, J Garzn-Ferreira, Seasonality in algal assemblages on upwelling-influenced coral reefs in the colombian caribbean, Bot. Mar., 45 (2002), 284–292. https://doi.org/10.1515/BOT.2002.028 doi: 10.1515/BOT.2002.028
|
Figures(7) / Tables(2)

Shengnan Zhao, Sanling Yuan. A coral reef benthic system with grazing intensity and immigrated macroalgae in deterministic and stochastic environments[J]. Mathematical Biosciences and Engineering, 2022, 19(4): 3449-3471. doi: 10.3934/mbe.2022159







 DownLoad:
DownLoad: