Citation: Zhiwei Huang, Gang Huang. Mathematical analysis on deterministic and stochastic lake ecosystem models[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 4723-4740. doi: 10.3934/mbe.2019237

| [1] | M. Rautio, H. Mariash and L. Forsstrom, Seasonal shifts between autochthonous and allochthon-ous carbon contributions to zooplankton diets in a subarctic lake, Limnol. Oceanogr., 56 (2011), 1513–1524. |
| [2] | S. E. Jones, C. T. Solomon and B. C. Weidel, Subsidy or subtraction: how do terrestrial inputs influence consumer production in lakes, Freshwater Rev., 5 (2012), 37–49. |
| [3] | K. A. Emery, G. M. Wilkinson, M. L. Pace, et al., Use of allochthonous resources by zooplankton in reservoirs, Hydrobiologia, 758 (2015), 257–269. |
| [4] | J. Cole, S. R. Carpenter, J. F. Kitchell, et al., Differential support of lake food webs by three types of terrestrial organic carbon, Ecol. Lett., 9 (2006), 558–568. |
| [5] | M. Berggren, S. E. Ziegler, N. F. Gelais, et al., Contrasting patterns of allochthony among three major groups of crustacean zooplankton in boreal and temperate lakes, Ecology, 95 (2014), 1947–1959. |
| [6] | M. T. Brett, M. Kainz, S. J. Taipale, et al., Phytoplankton, not allochthonous carbon, sustains herbivorous zooplankton production, Proc. Natl. Acad. Sci. USA, 21 (2009), 197–201. |
| [7] | M. Berggren, A. Bergstrom and J. Karlsson, Intraspecific autochthonous and allochthonous resource use by zooplankton in a humic lake during the transitions between winter, summer and fall, Plos One, 10 (2014), e0120575. |
| [8] | A. D. Persaud and P. J. Dillon, Differences in zooplankton feeding rates and isotopic signatures from three temperate lakes, Aquat. SCI., 73 (2011), 261–273. |
| [9] | N. D. Lewis, M. N. Breckels, M. Steinke, et al., Role of infochemical mediated zooplankton grazing in a phytoplankton competition model, Ecol. Complex., 16 (2013), 41–50. |
| [10] | Y. Lv and Y. Pei, Harvesting of a phytoplankton-zooplankton model, Nonlinear Anal. Real. World. Appl., 11 (2010), 3608–3619. |
| [11] | A. Sharma, A. K. Sharma and K. Agnihotri, Analysis of a toxin producing phytoplankton zooplankton interaction with Holling IV type scheme and time delay, Nonlinear Dynam., 81 (2015), 13–25. |
| [12] | J. E. Truscott and J. Brindley, Ocean plankton population as excitable media, B. Math. Biol., 56 (1994), 981–998. |
| [13] | Y. Pei, Y. Lv and C. Li, Evolutionary consequences of harvesting for a two-zooplankton one-phytoplankton system, Appl. Math. Model, 36 (2012), 1752–1765. |
| [14] | L. Qian, Q. Lu, Q. Meng et al., Dynamical behaviors of a prey-predator system with impulsive control, J. Math. Anal. Appl., 1 (2010), 345–356. |
| [15] | S. R. Jang and E. J. Allen, Deterministic and stochastic nutrient phytoplankton zooplankton models with periodic toxin producing phytoplankton, Appl. Math. Comput., 271 (2015), 52–67. |
| [16] | S. Ruan, Persistence and coexistence in zooplankton-phytoplankton nutrient models with instantaneous nutrient recycling, J. Math. Biol., 31 (1993), 633–654. |
| [17] | J. S. Wroblewski, S. L. Sarmiento and G. R. Flierl, An ocean basin scale model of plankton dynamics in the North Atlantic, solutions for the climatological oceanographic condition in May, Global Biogeochem. CY., 2 (1988), 199–218. |
| [18] | A. J. Tanentzap, B. W. Kielstra, G. M. Wilkinson, et al., Terrestrial support of lake food webs: synthesis reveals controls over cross-ecosystem resource use, Sci. Adv., 3 (2017), e1601765. |
| [19] | B. C. Mcmeans, A. Koussoroplis, M. T. Arts, et al., Terrestrial dissolved organic matter supports growth and reproduction of Daphnia magna when algae are limiting, J. Plankton. Res., 0 (2015), 1–9. |
| [20] | U. Forys, M. Qiao and A. Liu, Asymptotic dynamics of a deterministic and stochastic predator- prey model with disease in the prey species, Math. Method. Appl. Sci., 37 (2014), 306–320. |
| [21] | G. Ge, H. Wang and J. Xu, A stochastic analysis for a phytoplankton zooplankton model, JPCS, 96 (2008), 012168. |
| [22] | P. Ghosh, P. Das and D. Mukherjee, Persistence and stability of a seasonally perturbed three species stochastic model of salmonoid aquaculture, Differ. Equ. Dyn. Syst., 2016. Available from: https://doi.org/10.1007/s12591-016-0283-0. |
| [23] | C. Ji, D. Jiang and N. Shi, A note on a predator-prey model with modified Leslie-Gower and Holling-type II schemes with stochastic perturbation, J. Math. Anal. Appl., 377 (2011), 435–440. |
| [24] | J. Djordjevic, C. J. Silva and D. F. Torres, A stochastic SICA epidemic model for HIV transmission, Appl. Math. Lett., 84 (2018), 168–175. |
| [25] | R. Rajaji and M. Pitchaimani, Analysis of stochastic viral infection model with immune impairment, Int. J. Ap. Mat. Com-Pol., 3 (2017), 3561–3574. |
| [26] | F. C. Klebaner, Introduction to Stochastic Calculus with Applications, 2nd edition, World Scientific Publishing, London, 2005. |
| [27] | X. Mao, Stochastic Diferential Equations and Applications, 1nd edition, Horwood Publishing, Chichester, UK, 2008. |
| [28] | N. D. Lewis, M. N. Breckels, S. D. Archer, et al., Grazing-induced production of DMS can stabilize food-web dynamics and promote the formation of phytoplankton blooms in a multitrophic plankton model, Biogeochem., 110 (2012), 303–313. |
| [29] | D. J. Higham, An algorithmic introduction to numerical simulation of stochastic differential equations, SIAM Rev., 43 (2001), 525–546. |
| [30] | N. Dalal, D. Greenhalgh and X. Mao, A stochastic model for internal HIV dynamics, J. Math. Anal. Appl., 341 (2008), 1084–1101. |
| [31] | L. Shaikheta and A. Korobeinikov, Stability of a stochastic model for HIV-1 dynamics within a host, Appl. Anal., 6 (2016), 1228–1238. |
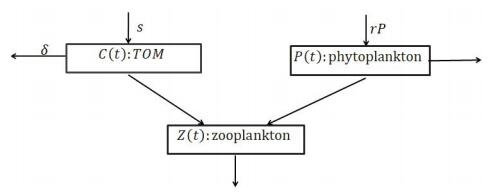
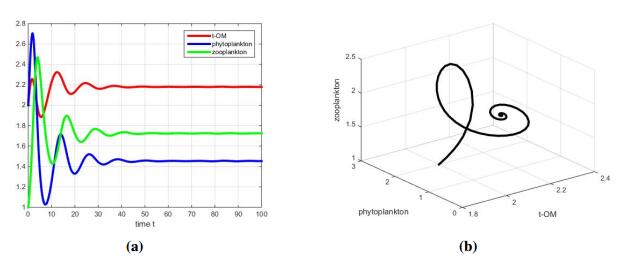
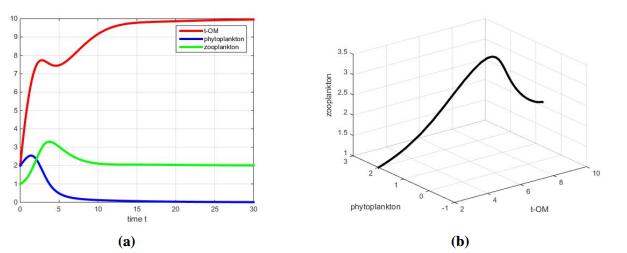
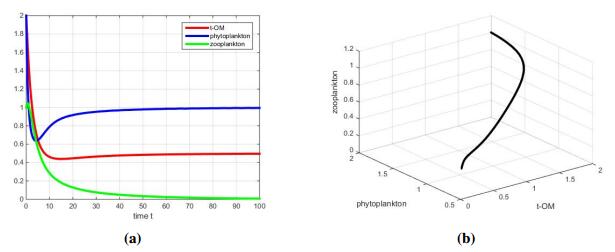
Figures(5) / Tables(2)

Zhiwei Huang, Gang Huang. Mathematical analysis on deterministic and stochastic lake ecosystem models[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 4723-4740. doi: 10.3934/mbe.2019237







 DownLoad:
DownLoad: