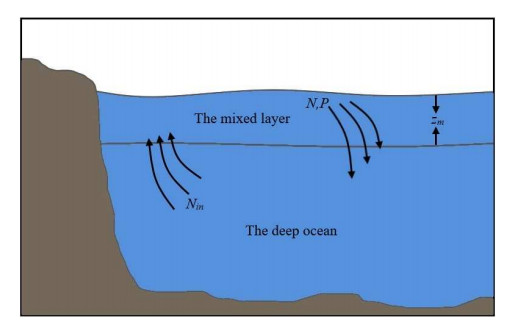
The variation of nutrient supply not only leads to the differences in the phytoplankton biomass and primary productivity but also induces the long-term phenotypic evolution of phytoplankton. It is widely accepted that marine phytoplankton follows Bergmann's Rule and becomes smaller with climate warming. Compared with the direct effect of increasing temperature, the indirect effect via nutrient supply is considered to be an important and dominant factor in the reduction of phytoplankton cell size. In this paper, a size-dependent nutrient-phytoplankton model is developed to explore the effects of nutrient supply on the evolutionary dynamics of functional traits associated with phytoplankton size. The ecological reproductive index is introduced to investigate the impacts of input nitrogen concentration and vertical mixing rate on the persistence of phytoplankton and the distribution of cell size. In addition, by applying the adaptive dynamics theory, we study the relationship between nutrient input and the evolutionary dynamics of phytoplankton. The results show that input nitrogen concentration and vertical mixing rate have significant effects on the cell size evolution of phytoplankton. Specifically, cell size tends to increase with the input nutrient concentration, as does the diversity of cell sizes. In addition, a single-peaked relationship between vertical mixing rate and cell size is observed. When the vertical mixing rate is too low or too high, only small individuals are dominant in the water column. When the vertical mixing rate is moderate, large individuals can coexist with small individuals, so the diversity of phytoplankton is elevated. We predict that reduced intensity of nutrient input due to climate warming will lead to a trend towards smaller cell size and will reduce the diversity of phytoplankton.
Citation: Lidan Liu, Meng Fan, Yun Kang. Effect of nutrient supply on cell size evolution of marine phytoplankton[J]. Mathematical Biosciences and Engineering, 2023, 20(3): 4714-4740. doi: 10.3934/mbe.2023218
The variation of nutrient supply not only leads to the differences in the phytoplankton biomass and primary productivity but also induces the long-term phenotypic evolution of phytoplankton. It is widely accepted that marine phytoplankton follows Bergmann's Rule and becomes smaller with climate warming. Compared with the direct effect of increasing temperature, the indirect effect via nutrient supply is considered to be an important and dominant factor in the reduction of phytoplankton cell size. In this paper, a size-dependent nutrient-phytoplankton model is developed to explore the effects of nutrient supply on the evolutionary dynamics of functional traits associated with phytoplankton size. The ecological reproductive index is introduced to investigate the impacts of input nitrogen concentration and vertical mixing rate on the persistence of phytoplankton and the distribution of cell size. In addition, by applying the adaptive dynamics theory, we study the relationship between nutrient input and the evolutionary dynamics of phytoplankton. The results show that input nitrogen concentration and vertical mixing rate have significant effects on the cell size evolution of phytoplankton. Specifically, cell size tends to increase with the input nutrient concentration, as does the diversity of cell sizes. In addition, a single-peaked relationship between vertical mixing rate and cell size is observed. When the vertical mixing rate is too low or too high, only small individuals are dominant in the water column. When the vertical mixing rate is moderate, large individuals can coexist with small individuals, so the diversity of phytoplankton is elevated. We predict that reduced intensity of nutrient input due to climate warming will lead to a trend towards smaller cell size and will reduce the diversity of phytoplankton.

| [1] |
J. Beardall, S. Stojkovic, S. Larsen, Living in a high CO$_2$ world: Impacts of global climate change on marine phytoplankton, Plant Ecol. Divers., 2 (2009), 191–205. https://doi.org/10.1080/17550870903271363 doi: 10.1080/17550870903271363

|
| [2] |
C. D. G. Harley, A. R. Hughes, K. M. Hultgren, B. G. Miner, C. J. B. Sorte, C. S. Thornber, et al., The impacts of climate change in coastal marine systems, Ecol. Lett., 9 (2006), 228–241. https://doi.org/10.1111/j.1461-0248.2005.00871.x doi: 10.1111/j.1461-0248.2005.00871.x
|
| [3] |
T. Zohary, G. Flaim, U. Sommer, Temperature and the size of freshwater phytoplankton, Hydrobiologia, 848 (2021), 143–155. https://doi.org/10.1007/s10750-020-04246-6 doi: 10.1007/s10750-020-04246-6
|
| [4] |
U. Sommer, K. H. Peter, S. Genitsaris, M. Moustaka-Gouni, Do marine phytoplankton follow Bergmann's rule sensu lato?, Biol. Rev., 92 (2017), 1011–1026. https://doi.org/10.1111/brv.12266 doi: 10.1111/brv.12266
|
| [5] |
E. Marañón, P. Cermeño, M. Latasa, R. D. Tadonléké, Resource supply alone explains the variability of marine phytoplankton size structure, Limnol. Oceanogr., 60 (2015), 1848–1854. https://doi.org/10.1002/lno.10138 doi: 10.1002/lno.10138
|
| [6] |
D. G. Boyce, M. R. Lewis, B. Worm, Global phytoplankton decline over the past century, Nature, 466 (2010), 591–596. https://doi.org/10.1038/nature09268 doi: 10.1038/nature09268
|
| [7] |
S. A. Henson, C. Beaulieu, T. Ilyina, J. G. John, M. Long, R. Séférian, et al., Rapid emergence of climate change in environmental drivers of marine ecosystems, Nat. Commun., 8 (2017), 1–9. https://doi.org/10.1038/ncomms14682 doi: 10.1038/ncomms14682
|
| [8] |
A. J. Irwin, Z. V. Finkel, O. M. E. Schofield, P. G. Falkowski, Scaling-up from nutrient physiology to the size-structure of phytoplankton communities, J. Plankton Res., 28 (2006), 459–471. https://doi.org/10.1093/plankt/fbi148 doi: 10.1093/plankt/fbi148
|
| [9] |
B. Bec, Y. Collos, A. Vaquer, D. Mouillot, P. Souchu, Growth rate peaks at intermediate cell size in marine photosynthetic picoeukaryotes, Limnol. Oceanogr., 53 (2008), 863–867. https://doi.org/10.4319/lo.2008.53.2.0863 doi: 10.4319/lo.2008.53.2.0863
|
| [10] |
A. E. Walsby, D. P. Holland, Sinking velocities of phytoplankton measured on a stable density gradient by laser scanning, J. R. Soc. Interface, 3 (2006), 429–439. https://doi.org/10.1098/rsif.2005.0106 doi: 10.1098/rsif.2005.0106
|
| [11] |
E. Litchman, C. Klausmeier, O. Schofield, P. Falkowski, The role of functional traits and trade-offs in structuring phytoplankton communities: Scaling from cellular to ecosystem level, Ecol. Lett., 10 (2007), 1170–1181. https://doi.org/10.1111/j.1461-0248.2007.01117.x doi: 10.1111/j.1461-0248.2007.01117.x
|
| [12] |
T. Key, A. McCarthy, D. A. Campbell, C. Six, S. Roy, Z. V. Finkel, Cell size trade-offs govern light exploitation strategies in marine phytoplankton, Environ. Microbiol., 12 (2010), 95–104. https://doi.org/10.1111/j.1462-2920.2009.02046.x doi: 10.1111/j.1462-2920.2009.02046.x
|
| [13] |
C. Serra-Pompei, G. I. Hagstrom, A. W. Visser, K. H. Andersen, Resource limitation determines temperature response of unicellular plankton communities, Limnol. Oceanogr., 64 (2019), 1627–1640. https://doi.org/10.1002/lno.11140 doi: 10.1002/lno.11140
|
| [14] |
K. H. Peter, U. Sommer, Phytoplankton cell size reduction in response to warming mediated by nutrient limitation, PLoS One, 8 (2013), 1–6. https://doi.org/10.1371/journal.pone.0071528 doi: 10.1371/journal.pone.0071528
|
| [15] |
Z. V. Finkel, J. Beardall, K. J. Flynn, A. Quigg, T. A. V. Rees, J. A. Raven, Phytoplankton in a changing world: cell size and elemental stoichiometry, J. Plankton Res., 32 (2010), 119–137. https://doi.org/10.1093/plankt/fbp098 doi: 10.1093/plankt/fbp098
|
| [16] |
A. Ryabov, O. Kerimoglu, E. Litchman, I. Olenina, L. Roselli, A. Basset, et al., Shape matters: the relationship between cell geometry and diversity in phytoplankton, Ecol. Lett., 24 (2021), 847-861. https://doi.org/10.1111/ele.13680 doi: 10.1111/ele.13680
|
| [17] |
E. Marañón, P. Cermeno, M. Latasa, R. D. Tadonléké, Temperature, resources, and phytoplankton size structure in the ocean, Limnol. Oceanogr., 57 (2012), 1266–1278. https://doi.org/10.4319/lo.2012.57.5.1266 doi: 10.4319/lo.2012.57.5.1266
|
| [18] |
D. B. Van de Waal, E. Litchman, Multiple global change stressor effects on phytoplankton nutrient acquisition in a future ocean, Philos. Trans. R. Soc. B-Biol. Sci., 375 (2020), 20190706. https://doi.org/10.1098/rstb.2019.0706 doi: 10.1098/rstb.2019.0706
|
| [19] |
E. Litchman, C. A. Klausmeier, K. Yoshiyama, Contrasting size evolution in marine and freshwater diatoms, Proc. Natl. Acad. Sci. U. S. A., 106 (2009), 2665–2670. https://doi.org/10.1073/pnas.0810891106 doi: 10.1073/pnas.0810891106
|
| [20] |
Z. Pu, M. H. Cortez, L. Jiang, Predator-prey coevolution drives productivity-richness relationships in planktonic systems, Am. Nat., 189 (2017), 28–42. https://doi.org/10.1086/689550 doi: 10.1086/689550
|
| [21] |
Y. Kang, J. H. Fewell, Co-evolutionary dynamics of a social parasite-host interaction model: Obligate versus facultative social parasitism, Nat. Resour. Model., 28 (2015), 398–455. https://doi.org/10.1111/nrm.12078 doi: 10.1111/nrm.12078
|
| [22] |
W. Zhang, S. Zhao, X. Meng, T. Zhang, Evolutionary analysis of adaptive dynamics model under variation of noise environment, Appl. Math. Model., 84 (2020), 222–239. https://doi.org/10.1016/j.apm.2020.03.045 doi: 10.1016/j.apm.2020.03.045
|
| [23] |
A. Li, X. Zou, Evolutionary analysis of adaptive dynamics model under variation of noise environment, Bull. Math. Biol., 83 (2021), 1-27. https://doi.org/10.1007/s11538-021-00893-5 doi: 10.1007/s11538-021-00893-5
|
| [24] |
P. Branco, M. Egas, S. R. Hall, J. Huisman, Why do phytoplankton evolve large size in response to grazing?, Am. Nat., 195 (2020), E20-E37. https://doi.org/ 10.1086/706251 doi: 10.1086/706251
|
| [25] |
H. Yang, Global dynamics of a diffusive phytoplankton-zooplankton model with toxic substances effect and delay, Math. Biosci. Eng., 19 (2022), 6712–6730. https://doi.org/10.3934/mbe.2022316 doi: 10.3934/mbe.2022316
|
| [26] |
L. Jiang, O. M. E. Schofield, P. G. Falkowski, Adaptive Evolution of Phytoplankton Cell Size, Am. Nat., 166 (2005), 496–505. https://doi.org/10.1086/444442 doi: 10.1086/444442
|
| [27] |
H. Wang, H. L. Smith, Y. Kuang, J. J. Elser, Dynamics of stoichiometric bacteria-algae interactions in the epilimnion, SIAM J. Appl. Math., 68 (2007), 503–522. https://doi.org/10.1137/060665919 doi: 10.1137/060665919
|
| [28] |
U. Dieckmann, R. Law, The dynamical theory of coevolution: A derivation from stochastic ecological processes, J. Math. Biol., 34 (1996), 579–612. https://doi.org/10.1007/BF02409751 doi: 10.1007/BF02409751
|
| [29] |
S. A. H. Geritz, É. Kisdi, G. Meszéna, J. A. J. Metz, Evolutionarily singular strategies and the adaptive growth and branching of the evolutionary tree, Evol. Ecol., 12 (1998), 35–57. https://doi.org/10.1023/A:1006554906681 doi: 10.1023/A:1006554906681
|
| [30] |
A. E. F. Prowe, M. Pahlow, A. Oschlies, Controls on the diversity-productivity relationship in a marine ecosystem model, Ecol. Model., 225 (2012), 167–176. https://doi.org/10.1016/j.ecolmodel.2011.11.018 doi: 10.1016/j.ecolmodel.2011.11.018
|
| [31] |
M. Adjou, J. Bendtsen, K. Richardson, Modeling the influence from ocean transport, mixing and grazing on phytoplankton diversity, Ecol. Model., 225 (2012), 19–27. https://doi.org/10.1016/j.ecolmodel.2011.11.005 doi: 10.1016/j.ecolmodel.2011.11.005
|
| [32] |
V. Huston, A theorem on average Liapunov functions, Monatsh. Math., 98 (1984), 267–275. https://doi.org/10.1007/BF01540776 doi: 10.1007/BF01540776
|








Figures(9) / Tables(1)

Lidan Liu, Meng Fan, Yun Kang. Effect of nutrient supply on cell size evolution of marine phytoplankton[J]. Mathematical Biosciences and Engineering, 2023, 20(3): 4714-4740. doi: 10.3934/mbe.2023218







 DownLoad:
DownLoad: