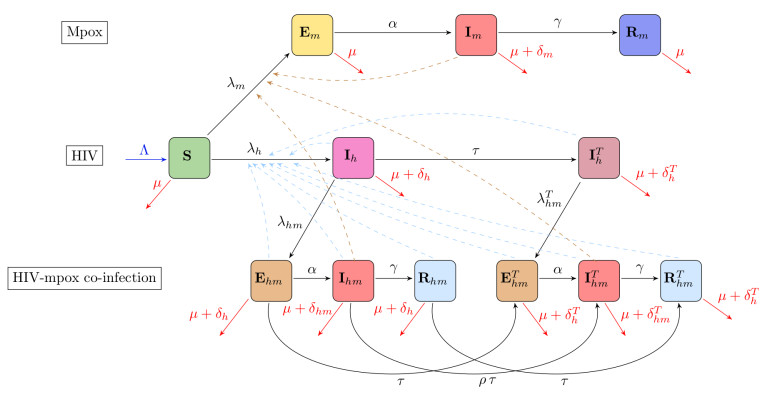
During the 2022 monkeypox (Mpox) outbreak in non-endemic countries, sexual transmission was identified as the dominant mode of transmission, and particularly affected the community of men who have sex with men (MSM). This community experienced the highest incidence of Mpox cases, exacerbating the public health burden they already face due to the disproportionate impact of HIV. Given the simultaneous spread of HIV and Mpox within the MSM community, it is crucial to understand how these diseases interact. Specifically, since HIV is endemic within this population, understanding its influence on the spread and control of Mpox is essential. In this study, we analyze a mechanistic mathematical model of Mpox to explore the potential impact of HIV on the dynamics of Mpox within the MSM community. The model considered in this work incorporates the transmission dynamics of the two diseases, including antiretroviral therapy (ART) for HIV. We assumed that HIV was already endemic in the population at the onset of the Mpox outbreak. Through our analysis, we derived the Mpox invasion reproduction number within an HIV-endemic setting and established the existence and local asymptotic stability of the Mpox-free equilibrium under these conditions. Furthermore, we demonstrated the existence and local asymptotic stability of an Mpox-endemic equilibrium in an HIV-endemic regime. Notably, our findings revealed that the model exhibits a backward bifurcation, a phenomenon that may not have occurred in the absence of HIV within the population. The public health significance of our results is that the presence of HIV in the MSM community could hinder efforts to control Mpox, allowing the disease to become endemic even when its invasion reproduction number is below one. Additionally, we found that Mpox might be more challenging to control in scenarios where HIV increases susceptibility to Mpox. Finally, consistent with previous studies, our analysis confirms that reducing sexual contact can be effective for controlling the spread of Mpox within the MSM community.
Citation: Andrew Omame, Sarafa A. Iyaniwura, Qing Han, Adeniyi Ebenezer, Nicola L. Bragazzi, Xiaoying Wang, Woldegebriel A. Woldegerima, Jude D. Kong. Dynamics of Mpox in an HIV endemic community: A mathematical modelling approach[J]. Mathematical Biosciences and Engineering, 2025, 22(2): 225-259. doi: 10.3934/mbe.2025010
During the 2022 monkeypox (Mpox) outbreak in non-endemic countries, sexual transmission was identified as the dominant mode of transmission, and particularly affected the community of men who have sex with men (MSM). This community experienced the highest incidence of Mpox cases, exacerbating the public health burden they already face due to the disproportionate impact of HIV. Given the simultaneous spread of HIV and Mpox within the MSM community, it is crucial to understand how these diseases interact. Specifically, since HIV is endemic within this population, understanding its influence on the spread and control of Mpox is essential. In this study, we analyze a mechanistic mathematical model of Mpox to explore the potential impact of HIV on the dynamics of Mpox within the MSM community. The model considered in this work incorporates the transmission dynamics of the two diseases, including antiretroviral therapy (ART) for HIV. We assumed that HIV was already endemic in the population at the onset of the Mpox outbreak. Through our analysis, we derived the Mpox invasion reproduction number within an HIV-endemic setting and established the existence and local asymptotic stability of the Mpox-free equilibrium under these conditions. Furthermore, we demonstrated the existence and local asymptotic stability of an Mpox-endemic equilibrium in an HIV-endemic regime. Notably, our findings revealed that the model exhibits a backward bifurcation, a phenomenon that may not have occurred in the absence of HIV within the population. The public health significance of our results is that the presence of HIV in the MSM community could hinder efforts to control Mpox, allowing the disease to become endemic even when its invasion reproduction number is below one. Additionally, we found that Mpox might be more challenging to control in scenarios where HIV increases susceptibility to Mpox. Finally, consistent with previous studies, our analysis confirms that reducing sexual contact can be effective for controlling the spread of Mpox within the MSM community.

| [1] |
M. Patel, M. Adnan, A. Aldarhami, A. S. Bazaid, N. H. Saeedi, A. A. Alkayyal, et al., Current insights into diagnosis, prevention strategies, treatment, therapeutic targets, and challenges of monkeypox(Mpox) infections in human populations, Life, 13 (2023), 249. https://doi.org/10.3390/life13010249 doi: 10.3390/life13010249

|
| [2] |
J. Lu, H. Xing, C.Wang, M. Tang, C.Wu, F. Ye, et al., Mpox (formerly monkeypox): Pathogenesis, prevention, and treatment, Signal Transduction Targeted Ther., 8 (2023), 458. https://doi.org/10.1038/s41392-023-01675-2 doi: 10.1038/s41392-023-01675-2
|
| [3] |
E. A. Falendysz, J. G. Lopera, T. E. Rocke, J. E. Osorio, Monkeypox virus in animals: Current knowledge of viral transmission and pathogenesis in wild animal reservoirs and captive animal models, Viruses, 15 (2003), 905. https://doi.org/10.3390/v15040905 doi: 10.3390/v15040905
|
| [4] |
N. Kumar, A. Acharya, H. E. Gendelman, S. N. Byrareddy, The 2022 outbreak and the pathobiology of the monkeypox virus, J. Autoimmun., 131 (2022), 102855. https://doi.org/10.1016/j.jaut.2022.102855 doi: 10.1016/j.jaut.2022.102855
|
| [5] | J. G. Breman, Kalisa-Ruti, M. V. Steniowski, E. Zanotto, A. I. Gromyko, I. Arita, Human monkeypox, 1970–1979, 1980. Available from: https://iris.who.int/handle/10665/67095. |
| [6] |
D. L. Heymann, M. Szczeniowski, K. Esteves, Re-emergence of monkeypox in africa: A review of the past six years, British Med. Bull., 54 (1998), 693–702. https://doi.org/10.1093/oxfordjournals.bmb.a011720 doi: 10.1093/oxfordjournals.bmb.a011720
|
| [7] | H. Nilasari, M. Trifitriana, R. Anadya, L. Ameline, D. Pratiwi, The evolving monkeypox outbreak amongst homosexual and bisexual transmission: A systematic review, J. Pakistan Assoc. Dermat., 34 (2024), 243–254. https://www.jpad.com.pk/index.php/jpad/article/view/2502 |
| [8] |
A. M. McCollum, V. Shelus, A. Hill, T. Traore, B. Onoja, Y. Nakazawa, et al., Epidemiology of human mpox—worldwide, 2018–2021, MMWR Morb. Mortal. Wkly. Rep., 72 (2023), 68–72. http://dx.doi.org/10.15585/mmwr.mm7203a4 doi: 10.15585/mmwr.mm7203a4
|
| [9] |
M. G. Reynolds, I. K. Damon, Outbreaks of human monkeypox after cessation of smallpox vaccination, Trends Microbiol., 20 (2012), 80–87. https://doi.org/10.1016/j.tim.2011.12.001 doi: 10.1016/j.tim.2011.12.001
|
| [10] |
G. M. Zaucha, P. B. Jahrling, T. W. Geisbert, J. R. Swearengen, L. Hensley, The pathology of experimental aerosolized monkeypox virus infection in cynomolgus monkeys (macaca fascicularis), Lab. Invest., 81 (2001), 1581–1600. https://doi.org/10.1038/labinvest.3780373 doi: 10.1038/labinvest.3780373
|
| [11] | J. Kaler, A. Hussain, G. Flores, S. Kheiri, D. Desrosiers, Monkeypox: A comprehensive review of transmission, pathogenesis, and manifestation, Cureus, 14 (2022). https://doi.org/10.7759/cureus.26531 |
| [12] |
P. Kumar, B. Chaudhary, N. Yadav, S. Devi, A. Pareek, S. Alla, et al., Recent advances in research and management of human monkeypox virus: An emerging global health threat, Viruses, 15 (2023), 937. https://doi.org/10.3390/v15040937 doi: 10.3390/v15040937
|
| [13] |
K. Mellou, K. Tryfinopoulou, S. Pappa, K. Gkolfinopoulou, S. Papanikou, G. Papadopoulou, et. al., Overview of Mpox outbreak in Greece in 2022–2023: Is it over, Viruses, 15 (2023), 1384. https://doi.org/10.3390/v15061384 doi: 10.3390/v15061384
|
| [14] |
S. T. Al Awaidy, F. Khamis, M. Sallam, R. M. Ghazy, H. Zaraket, Monkeypox (Mpox) outbreak: More queries posed as cases soar globally, Sultan Qaboos Univ. Med. J., 23 (2023), 1–4. https://doi.org/10.18295/squmj.8.2022.046 doi: 10.18295/squmj.8.2022.046
|
| [15] |
L. T. Allan-Blitz, M. Gandhi, P. Adamson, I. Park, G. Bolan, J. D. Klausner, A position statement on mpox as a sexually transmitted disease, Clin. Infect. Dis., 76 (2013), 1508–1512. https://doi.org/10.1093/cid/ciac960 doi: 10.1093/cid/ciac960
|
| [16] | RR Assessment, Monkeypox multi-country outbreak, Eur. Cent. Dis. Prev. Control, 2022 (2022). |
| [17] |
B. Ortiz-Saavedra, E. S. Montes-Madariaga, C. Cabanillas-Ramirez, N. Alva, A. Ricardo-Martinez, D. A. Leon-Figueroa, et al., Epidemiologic situation of HIV and monkeypox coinfection: A systematic review, Vaccines, 11 (2023), 246. https://doi.org/10.3390/vaccines11020246 doi: 10.3390/vaccines11020246
|
| [18] |
S. Cahill, Lessons learned from the us public health response to the 2022 mpox outbreak, LGBT Health, 10 (2023), 489–495. https://doi.org/10.1089/lgbt.2022.0274 doi: 10.1089/lgbt.2022.0274
|
| [19] |
K. K. Kota, J. Hong, C. Zelaya, A. P. Riser, A. Rodriguez, D. L. Weller, et al., Racial and ethnic disparities in mpox cases and vaccination among adult males—united states, may-december 2022, MMWR Morb. Mortal. Wkly. Rep., 72 (2023), 398–403. http://dx.doi.org/10.15585/mmwr.mm7215a4 doi: 10.15585/mmwr.mm7215a4
|
| [20] |
H. Sachdeva, R. Shahin, S. Ota, S. Isabel, C. S. Mangat, R. Stuart, et al., Preparing for mpox resurgence: Surveillance lessons from outbreaks in toronto, canada, J. Infect. Dis., 229 (2024), S305–S312. https://doi.org/10.1093/infdis/jiad533 doi: 10.1093/infdis/jiad533
|
| [21] |
A. Zebardast, T. Latifi, N. Jandaghi, M. G. Barzoki, S. Malekshahi, Plausible reasons for the resurgence of mpox (formerly monkeypox): An overview, Trop. Dis., Travel Med. Vaccines, 9 (2023), 23 https://doi.org/10.1186/s40794-023-00209-6 doi: 10.1186/s40794-023-00209-6
|
| [22] |
S. Mohanto, M. Faiyazuddin, A. D. Gholap, D. Jogi, A. Bhunia, K. Subbaram, et al., Addressing the resurgence of global monkeypox (Mpox) through advanced drug delivery platforms, Travel Med. Infect. Dis., 56 (2023), 102636. https://doi.org/10.1016/j.tmaid.2023.102636 doi: 10.1016/j.tmaid.2023.102636
|
| [23] |
L. M. Masirika, A. Kumar, M. Dutt, A. T. Ostadgavahi, B. Hewins, N. M. Bubala, et. al., Complete genome sequencing, annotation, and mutational profiling of the novel clade I human Mpox virus, Kamituga strain, J. Infect. Dev. Ctries, 18 (2024), 600–608. https://doi.org/10.3855/jidc.20136 doi: 10.3855/jidc.20136
|
| [24] |
E. Alakunle, D. Kolawole, D. Diaz-Canova, F. Alele, O. Adegboye, U. Moens, et al., A comprehensive review of monkeypox virus and Mpox characteristics, Front. Cell. Infect. Microbiol., 14 (2024), 1360586. https://doi.org/10.3389/fcimb.2024.1360586 doi: 10.3389/fcimb.2024.1360586
|
| [25] | L. Taylor, Mpox in Africa: WHO and Africa CDC consider declaring public health emergency as cases spike, British Med. J., 386 (2024). https://doi.org/10.1136/bmj.q1795 |
| [26] | World Health Organization, WHO director-general declares Mpox outbreak a public health emergency of international concern, 2024. |
| [27] | Kaiser Family Foundation, Black Americans and HIV/AIDS: The basics, 2023. |
| [28] | Global statistics, 2024. Available from: https://www.hiv.gov/hiv-basics/overview/data-and-trends/global-statistics. |
| [29] | Global HIV AIDS statistics—Fact sheet, 2024. Available from: https://www.unaids.org/en/resources/fact-sheet. |
| [30] | H. Swinkels, A. J. Vaillant, A. Nguyen, P. Gulick. HIV and AIDS. StatPearls, (2024). Available from: https://www.ncbi.nlm.nih.gov/books/NBK534860/. |
| [31] | Centers for Disease Control and Prevention, How HIV spreads, 2023. Available from: https://www.cdc.gov/hiv/causes/index.html. |
| [32] |
Q. Tollett, S. Safdar, A. B. Gumel, Dynamics of a two-group model for assessing the impacts of pre-exposure prophylaxis, testing and risk behaviour change on the spread and control of HIV/AIDS in an MSM population, Infect. Dis. Modell., 9 (2024), 103–127. https://doi.org/10.1016/j.idm.2023.11.004 doi: 10.1016/j.idm.2023.11.004
|
| [33] |
D. Patel, C. H. Johnson, A. Krueger, B. Maciak, L. Belcher, N. Harris, et al., Trends in HIV testing among US adults, aged 18-64 years, 2011–2017, AIDS Behav., 24 (2020), 532–539. https://doi.org/10.1007/s10461-019-02689-0 doi: 10.1007/s10461-019-02689-0
|
| [34] |
S. J. Challacombe, Global inequalities in HIV infection, Oral Dis., 26 (2020), 16–21. https://doi.org/10.1111/odi.13386 doi: 10.1111/odi.13386
|
| [35] |
A. T. Boyd, I. Oboho, H. Paulin, H. Ali, C. Godfrey, A. Date, et al., Addressing advanced HIV disease and mortality in global HIV programming, AIDS Res. Ther., 17 (2020), 1–7. https://doi.org/10.1186/s12981-020-00296-x doi: 10.1186/s12981-020-00296-x
|
| [36] | Statista, Death rate for human immunodeficiency virus (HIV) in Canada from 2000 to 2022, 2024. |
| [37] | Common conditions and diseases in HIV-positive men who have sex with men, 2024. Available from: https://www.ohtn.on.ca/rapid-response-82-conditions-diseases-hiv-positive-gay-men. |
| [38] |
O. Mitja, A. Alemany, M. Marks, J. Mora, J. Rodríguez-Aldama, M. Silva, et. al., Mpox in people with advanced HIV infection: A global case series, The Lancet, 401 (2023), 939–949. https://doi.org/10.1016/S0140-6736(23)00273-8 doi: 10.1016/S0140-6736(23)00273-8
|
| [39] | O. Mitja, D. Ogoina, B. K. Titanji, C. Galvan, J-J Muyembe, M. Marks, et al., Monkeypox, The Lancet, 401 (2023), 60–74. https://doi.org/10.1016/S0140-6736(22)02075-X |
| [40] | Centers for Disease Control and Prevention, CDC health advisory; severe manifestations of monkeypox among people who are immunocompromised due to HIV or other conditions, 2023. Available from: https://emergency.cdc.gov/han/2022/pdf/CDC_HAN_475.pdf. |
| [41] | Centers for disease control and prevention, Clinical considerations for treatment and prophylaxis of mpox infection in people who are immunocompromised, 2023. Available from: https://www.cdc.gov/poxvirus/mpox/clinicians/people-with-HIV.html. |
| [42] |
J. Elford, G. Hart, If HIV prevention works, why are rates of high-risk sexual behavior increasing among MSM, AIDS Educ. Prev., 15 (2003), 294–308. https://doi.org/10.1521/aeap.15.5.294.23825 doi: 10.1521/aeap.15.5.294.23825
|
| [43] |
W. Thienkrua, F. van Griensven, P. A. Mock, E. F. Dunne, B. Raengsakulrach, W. Wimonsate, et al., Young men who have sex with men at high risk for HIV, Bangkok MSM cohort study, Thailand 2006–2014, AIDS Behav., 22 (2018), 2137–2146. https://doi.org/10.1007/s10461-017-1963-7 doi: 10.1007/s10461-017-1963-7
|
| [44] |
S. K. Sulaiman, F. Isma'il Tsiga-Ahmed, M. S. Musa, B. T. Makama, A. K. Sulaiman, T. B. Abdulaziz, Global prevalence and correlates of Mpox vaccine acceptance and uptake: A systematic review and meta-analysis, Commun. Med., 4 (2024), 136. https://doi.org/10.1038/s43856-024-00564-1 doi: 10.1038/s43856-024-00564-1
|
| [45] |
R. A. Ghaffar, S. Shahnoor, M. Farooq, Increased prevalence of HIV among monkeypox patients–an alarming update, New Microbes New Infect., 49-50 (2022), 101039. https://doi.org/10.1016/j.nmni.2022.101039 doi: 10.1016/j.nmni.2022.101039
|
| [46] |
J. Kowalski, I. Cielniak, E. Garbacz-Łagożna, G. Cholewińska-Szymańska, M. Parczewski, Comparison of clinical course of mpox among HIV-negative and HIV-positive patients: A 2022 cohort of hospitalized patients in central Europe, J. Med. Virol., 95 (2023), e29172. https://doi.org/10.1002/jmv.29172 doi: 10.1002/jmv.29172
|
| [47] | C. P. Bhunu, S. Mushayabasa, J. M. Hyman, Modelling HIV/AIDS and monkeypox co-infection. Appl. Math. Comput., 218 (2012), 9504–9518. https://doi.org/10.1016/j.amc.2012.03.042 |
| [48] |
O. I. Marcus, A. Augustine, T. Jonathan, A co-infection model for monkeypox and HIV/AIDS: Sensitivity and bifurcation analyses, J. Scient. Res. Rep., 30 (2024), 351–368. https://doi.org/10.9734/jsrr/2024/v30i51951 doi: 10.9734/jsrr/2024/v30i51951
|
| [49] | O. O. Peace, O. A. Godwin, B. Bolaji, A compartmental deterministic epidemiological model with non-linear differential equations for analyzing the co-infection dynamics between COVID-19, HIV, and monkeypox diseases, Healthcare Anal., (2024), 100311. https://doi.org/10.1016/j.health.2024.100311 |
| [50] |
A. Omame, Q. Han, S. A. Iyaniwura, E. Adeniyi, N. L. Bragazzi, X. Wang, et al., Understanding the impact of HIV on mpox transmission in an MSM population: A mathematical modeling study, Infect. Dis. Modell., 9 (2024), 1117–1137. https://doi.org/10.1016/j.idm.2024.05.008 doi: 10.1016/j.idm.2024.05.008
|
| [51] |
M. S. Cohen, Y. Q. Chen, M. McCauley, T. Gamble, M. C. Hosseinipour, N. Kumarasamy, et al., Prevention of HIV-1 infection with early antiretroviral therapy, N. Engl. J. Med., 365 (2011), 493–505. https://doi.org/10.1056/NEJMoa1105243 doi: 10.1056/NEJMoa1105243
|
| [52] |
R. W. Eisinger, C. W. Dieffenbach, A. S. Fauci, HIV viral load and transmissibility of HIV infection, JAMA, 321 (2019), 451–452. https://doi.org/10.1001/jama.2018.21167 doi: 10.1001/jama.2018.21167
|
| [53] | World Bank Group, Population, total-Canada, 2023. Available from: https://data.worldbank.org/indicator/SP.POP.TOTL?locations = CA. |
| [54] |
N. L. Bragazzi, Q. Han, S. A. Iyaniwura, A. Omame, A. Shausan, X. Wang, et al., Adaptive changes in sexual behavior in the high-risk population in response to human monkeypox transmission in canada can help control the outbreak: Insights from a two-group, two-route epidemic model, J. Med. Virol., 95 (2023), e28575. https://doi.org/10.1002/jmv.28575 doi: 10.1002/jmv.28575
|
| [55] | M. Xiridou, F. Miura, P. Adam, E. Op de Coul, J. de Wit, J. Wallinga, The fading of the mpox outbreak among men who have sex with men: A mathematical modelling study, J. Infect. Dis., (2023), e121–e130. https://doi.org/10.1093/infdis/jiad414 |
| [56] |
E. Beer, V. B. Rao, A systematic review of the epidemiology of human monkeypox outbreaks and implications for outbreak strategy, PLoS Negl. Trop. Dis., 13 (2019), e0007791. https://doi.org/10.1371/journal.pntd.0007791 doi: 10.1371/journal.pntd.0007791
|
| [57] |
F. Miura, C. E. van Ewijk, J. A. Backer, M. Xiridou, E. Franz, E. Op de Coul, et al., Estimated incubation period for monkeypox cases confirmed in the Netherlands, May 2022, Eurosurveillance, 27 (2022), 2200448. https://doi.org/10.2807/1560-7917.ES.2022.27.24.2200448 doi: 10.2807/1560-7917.ES.2022.27.24.2200448
|
| [58] | I. H. Spicknall, E. D. Pollock, P. A. Clay, A. M. Oster, K. Charniga, N. Masters, et al., Modeling the impact of sexual networks in the transmission of monkeypox virus among gay, bisexual, and other men who have sex with men—United States, 2022, Morb. Mortal. Wkly. Rep., 71 (2022), 1131–1135. |
| [59] |
V. D. Lima, J. Zhu, K. G. Card, N. J. Lachowsky, G. Chowell-Puente, Z. Wu, et al., Can the combination of TasP and PreP eliminate HIV among MSM in British Columbia, Canada, Epidemics, 35 (2021), 100461. https://doi.org/10.1016/j.epidem.2021.100461 doi: 10.1016/j.epidem.2021.100461
|
| [60] |
Y. Shen, C. Zhang, M. A. Valimaki, H. Qian, L. Mohammadi, Y. Chi, et al., Why do men who have sex with men practice condomless sex? A systematic review and meta-synthesis, BMC Infect. Dis., 22 (2022), 850. https://doi.org/10.1186/s12879-022-07843-z doi: 10.1186/s12879-022-07843-z
|
| [61] |
Z. Mukandavire, K. Bowa, W. Garira, Modelling circumcision and condom use as HIV/AIDS preventive control strategies, Math. Comput. Modell., 46 (2007), 1353–1372. https://doi.org/10.1016/j.mcm.2007.01.001 doi: 10.1016/j.mcm.2007.01.001
|
| [62] |
D. R. MacFadden, D. H. Tan, S. Mishra, Optimizing HIV pre-exposure prophylaxis implementation among men who have sex with men in a large urban centre: A dynamic modelling study, J. Int. AIDS Soc., 19 (2016), 20791. https://doi.org/10.7448/IAS.19.1.20791 doi: 10.7448/IAS.19.1.20791
|
| [63] | Centers for Disease Control and Prevention, Impact of monkeypox outbreak on select behaviors, 2023. Available from: https://www.cdc.gov/poxvirus/monkeypox/response/2022/amis-select-behaviors.html. |
| [64] |
P. van den Driessche, J. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180 (2002), 29–48. https://doi.org/10.1016/S0025-5564(02)00108-6 doi: 10.1016/S0025-5564(02)00108-6
|
| [65] | E. J. Routh, A treatise on the stability of a given state of motion, particularly steady motion: being the essay to which the Adams prize was adjudged in 1877, in the University of Cambridge. Macmillan and Company, 1877. |
| [66] | F. J. Kraus, M. Mansour, M. Sebek, Hurwitz matrix for polynomial matrices, in Stability Theory: Hurwitz Centenary Conference Centro Stefano Franscini, Ascona, 1995, Springer, (1996), 67–74. |
| [67] |
O. C. Collins, K. J. Duffy, Dynamics and control of mpox disease using two modelling approaches, Model. Earth Syst. Environ., 10 (2024), 1657–1669. https://doi.org/10.1007/s40808-023-01862-8 doi: 10.1007/s40808-023-01862-8
|
| [68] |
M. M. Al-Shomrani, S. S. Musa, A. Yusuf, Unfolding the transmission dynamics of monkeypox virus: An epidemiological modelling analysis, Mathematics, 11 (2023), 1121. https://doi.org/10.3390/math11051121 doi: 10.3390/math11051121
|
| [69] |
L. Wang, Global dynamical analysis of hiv models with treatments, Int. J. Bifurcation Chaos, 22 (2012), 1250227. https://doi.org/10.1142/S0218127412502276 doi: 10.1142/S0218127412502276
|
| [70] | Z. Mukandavire, P. Das, C. Chiyaka, F. Nyabadza, Global analysis of an HIV/AIDS epidemic model, World J. modelling and simulation, 6 (2010), 231–240. |
| [71] | Public Health Agency of Canada, Epidemiological summary report: 2022-23 Mpox outbreak in Canada, 2024. |
| [72] | E. Slaymaker, J. Todd, M. Marston, C. Calvert, D. Michael, J. Nakiyingi-Miiro, et al., How have ART treatment programmes changed the patterns of excess mortality in people living with HIV? estimates from four countries in East and Southern Africa, Global Health Action, 7 (2014). https://doi.org/10.3402/gha.v7.22789 |
| [73] | MathWorks, Matlab r2024a, 2024. Available from: https://matlab.mathworks.com. |
| [74] |
C. Castillo-Chavez, B. Song, Dynamical models of tuberculosis and their applications, Math. Biosci. Eng., 1 (2004), 361–404. https://doi.org/10.3934/mbe.2004.1.361 doi: 10.3934/mbe.2004.1.361
|
| [75] |
A. B. Gumel, Causes of backward bifurcations in some epidemiological models, J. Math. Anal. Appl., 395 (2012), 355–365. https://doi.org/10.1016/j.jmaa.2012.04.077 doi: 10.1016/j.jmaa.2012.04.077
|
| [76] |
B. Anderson, J. Jackson, M. Sitharam, Descartes' rule of signs revisited, Am. Math, Mon., 105 (1998), 447–451. https://doi.org/10.1080/00029890.1998.12004907 doi: 10.1080/00029890.1998.12004907
|
| [77] |
B. J. Turner, Adherence to antiretroviral therapy by human immunodeficiency virus—infected patients, J. Infect. Dis., 185 (2002), S143–S151. https://doi.org/10.1038/s41598-018-21081-x doi: 10.1038/s41598-018-21081-x
|
| [78] |
C. C. Hoff, D. Chakravarty, S. C. Beougher, T. B. Neilands, L. A. Darbes, Relationship characteristics associated with sexual risk behavior among MSM in committed relationships, AIDS Patient Care STDs, 26 (2012), 738–745. https://doi.org/10.1089/apc.2012.0198 doi: 10.1089/apc.2012.0198
|
| [79] |
B. R. Simon Rosser, K. J. Horvath, L. A. Hatfield, J. L. Peterson, S. Jacoby, A. Stately, et al., Predictors of HIV disclosure to secondary partners and sexual risk behavior among a high-risk sample of HIV-positive MSM: Results from six epicenters in the US, AIDS care, 20 (2008), 925–930. https://doi.org/10.1080/09540120701767265 doi: 10.1080/09540120701767265
|
| [80] |
D. Ogoina, M. Iroezindu, H. I. James, R. Oladokun, A. Yinka-Ogunleye, P. Wakama, et al., Clinical course and outcome of human monkeypox in Nigeria, Clin. Infect. Dis., 71 (2020), e210–e214. https://doi.org/10.1093/cid/ciaa143 doi: 10.1093/cid/ciaa143
|
| [81] |
M. A. Zinnah, M. B. Uddin, T. Hasan, S. Das, F. Khatun, M. H. Hasan, et al., The re-emergence of mpox: Old illness, modern challenges, Biomedicines, 12 (2024), 1457. https://doi.org/10.3390/biomedicines12071457 doi: 10.3390/biomedicines12071457
|
| [82] |
E. H. Vakaniaki, C. Kacita, E. Kinganda-Lusamaki, A. O'Toole, T. Wawina-Bokalanga, D. Mukadi-Bamuleka, et al., Sustained human outbreak of a new MPXV clade I lineage in the Eastern Democratic Republic of the Congo, Nat. Med., 30 (2024), 2791–2795. https://doi.org/10.1038/s41591-024-03130-3 doi: 10.1038/s41591-024-03130-3
|
| [83] |
D. A. Schwartz, High rates of miscarriage and stillbirth among pregnant women with clade I Mpox(monkeypox) are confirmed during 2023–2024 DR Congo outbreak in South Kivu Province, Viruses, 16 (2024), 1123. https://doi.org/10.3390/v16071123 doi: 10.3390/v16071123
|
| [84] |
L. Zhang, M. ur Rahman, M. Arfan, A. Ali, Investigation of mathematical model of transmission co-infection TB in HIV community with a non-singular kernel, Results Phys., 28 (2021), 104559. https://doi.org/10.1016/j.rinp.2021.104559 doi: 10.1016/j.rinp.2021.104559
|
| [85] |
N. Mohankumar, L. Rajagopal, J. J. Nieto, Optimal control for co-infection with COVID-19-associated pulmonary aspergillosis in icu patients with environmental contamination, Math. Biosci. Eng., 20 (2023), 9861–9875. https://doi.org/10.3934/mbe.2023432 doi: 10.3934/mbe.2023432
|
| [86] |
C. J. Silva, C. Cruz, D. F. M. Torres, A. P. Munuzuri, A. Carballosa, I. Area, et al., Optimal control of the COVID-19 pandemic: Controlled sanitary deconfinement in Portugal, Sci. Rep., 11 (2021), 3451. https://doi.org/10.1038/s41598-021-83075-6 doi: 10.1038/s41598-021-83075-6
|
| [87] |
B. Li, Z. Eskandari, Dynamical analysis of a discrete-time SIR epidemic model, J. Franklin Inst., 360 (2023), 7989–8007. https://doi.org/10.1016/j.jfranklin.2023.06.006 doi: 10.1016/j.jfranklin.2023.06.006
|
| [88] |
C. Mitchell, C. Kribs, Invasion reproductive numbers for periodic epidemic models, Infect. Dis. Modell., 4 (2019), 124–141. https://doi.org/10.1016/j.idm.2019.04.002 doi: 10.1016/j.idm.2019.04.002
|





Figures(6) / Tables(1)

Andrew Omame, Sarafa A. Iyaniwura, Qing Han, Adeniyi Ebenezer, Nicola L. Bragazzi, Xiaoying Wang, Woldegebriel A. Woldegerima, Jude D. Kong. Dynamics of Mpox in an HIV endemic community: A mathematical modelling approach[J]. Mathematical Biosciences and Engineering, 2025, 22(2): 225-259. doi: 10.3934/mbe.2025010







 DownLoad:
DownLoad: