Citation: Ya-Dong Zhang, Hai-Feng Huo, Hong Xiang. Dynamics of tuberculosis with fast and slow progression and media coverage[J]. Mathematical Biosciences and Engineering, 2019, 16(3): 1150-1170. doi: 10.3934/mbe.2019055

| [1] | World Health Organization, Global tuberculosis report 2017, Available from: https://www. who.int/tb/publications/global_report/en. |
| [2] | D.P. Gao and N.J. Huang, Optimal control analysis of a tuberculosis model, Appl. Math. Model., 58 (2018), 47–64. |
| [3] | S. Kim, A.A. Reyes and E. Jung, Mathematical model and intervention strategies for mitigating tuberculosis in the Philippines, J. Theor. Biol., 443 (2018), 100–112. |
| [4] | C.J. Silva, H. Maurer and D.F. Torres, Optimal control of a tuberculosis model with state and control delays, Math. Biosci. Eng., 14 (2016), 321–337. |
| [5] | H.F. Huo, S.J. Dang and Y.N. Li, Stability of a two-strain tuberculosis model with general contact rate, Abstr. Appl. Anal., 2010 (2010), 1–31. |
| [6] | H.F. Huo and M.X. Zou, Modelling effects of treatment at home on tuberculosis transmission dynamics, Appl. Math. Model., 40 (2016), 9474–9484. |
| [7] | H.F. Huo and L.X. Feng, Global stability for an HIV/AIDS epidemic model with different latent stages and treatment, Appl. Math. Model., 37 (2013), 1480–1489. |
| [8] | C.C. Mccluskey, Lyapunov functions for tuberculosis models with fast and slow progression, Math. Biosci. Eng., 3 (2006), 603–614. |
| [9] | T. Bergeb, S. Bowonga and J.M. Lubuma, Global stability of a two-patch cholera model with fast and slow transmissions, Math. Comput. Simulat., 133 (2017), 124–164. |
| [10] | B. Song, C. Castillo-Chavez and J.P. Aparicio, Tuberculosis models with fast and slow dynamics: the role of close and casual contacts, Math. Biosci., 180 (2002), 187–205. |
| [11] | J. Li, Y.L. Zhao and S.M. Li, Fast and slow dynamics of malaria model with relapse, Math. Biosci., 246 (2013), 94–104. |
| [12] | J.A. Cui, X. Tao and H.P. Zhu, An SIS infection model incorporating media coverage, Rocky Mt. J. Math., 38 (2008), 1323–1334. |
| [13] | H.F. Huo and X.M. Zhang, Complex dynamics in an alcoholism model with the impact of twitter, Math. Biosci., 281 (2016), 24–35. |
| [14] | H.F. Huo, P. Yang and H. Xiang, Stability and bifurcation for an SEIS epidemic model with the impact of media, Physica A, 490 (2018), 702–720. |
| [15] | J.G. Cui, Y.H. Sun and H.P. Zhu, The impact of media on the control of infectious diseases, J. Differ. Equations, 20 (2008), 31–53. |
| [16] | H. Xiang, Y.Y. Wang and H.F. Huo, Analysis of the binge drinking models with demographics and nonlinear infectivity on networks, J. Appl. Anal. Comput., 8 (2018), 1535–1554. |
| [17] | H.F. Huo, F.F. Cui and H. Xiang, Dynamics of an SAITS alcoholism model on unweighted and weighted networks, Physica A, 496 (2018), 249–262. |
| [18] | Y.L. Cai, J.J. Jiao, Z.J. Gui, Y.T. Liu and W.M. Wang, Environmental variability in a stochastic epidemic model, Appl. Math. Comput., 329 (2018), 210–226. |
| [19] | Z. Du and Z. Feng, Existence and asymptotic behaviors of traveling waves of a modified vectordisease model, Commun. Pur. Appl. Anal., 17 (2018), 1899–1920. |
| [20] | X.B. Zhang, Q.H. Shi, S.H. Ma, H.F. Huo and D.G. Li, Dynamic behavior of a stochastic SIQS epidemic model with levy jumps, Nonlinear Dynam., 93 (2018), 1481–1493. |
| [21] | W.M. Wang, Y.L. Cai, Z.Q. Ding and Z.J. Gui, A stochastic differential equation SIS epidemic model incorporating Ornstein-Uhlenbeck process, Physica A, 509 (2018), 921–936. |
| [22] | P.V. Driessche and J.Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180 (2002), 29–48. |
| [23] | J.P. LaSalle, The Stability of Dynamical Systems, Regional Conference Series in Applied Mathmatics, Society for Industrial and Applied Mathematics, Berlin, New Jersey, 1976. |
| [24] | J.D. Murray, Mathematical Biology, Springer-Verlag, Berlin, 1998. |
| [25] | C. Castillo-Chavez and B. Song, Dynamical models of tuberculosis and their applications, Math. Biosci. Eng., 1 (2004), 361–401. |
| [26] | K.A. Pawelek, A. Oeldorfhirsch and L. Rong, Modeling the impact of twitter on influenza epidemics, Math. Biosci. Eng., 11 (2014), 1337–1356. |
| [27] | K. Styblo, J. Meijer and I. Sutherland, The transmission of Tubercle Bacilli: its trend in a human population, Bull. World Health Organ., 41 (1969), 137–178. |
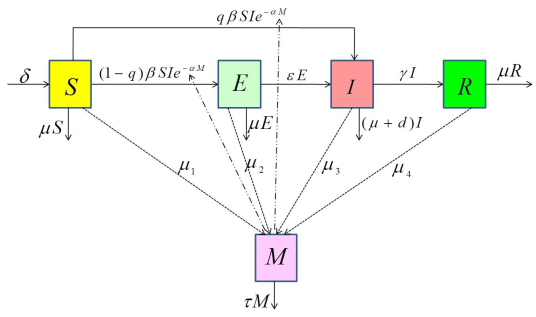
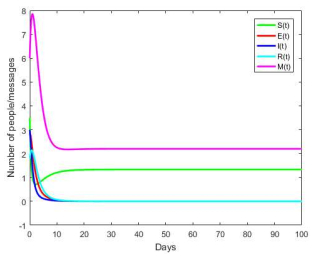
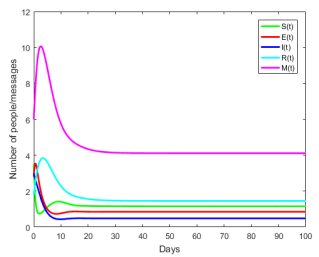
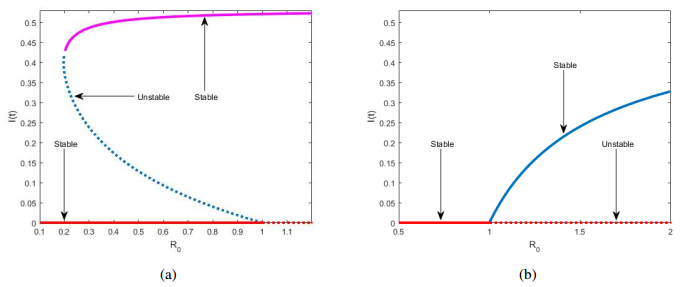
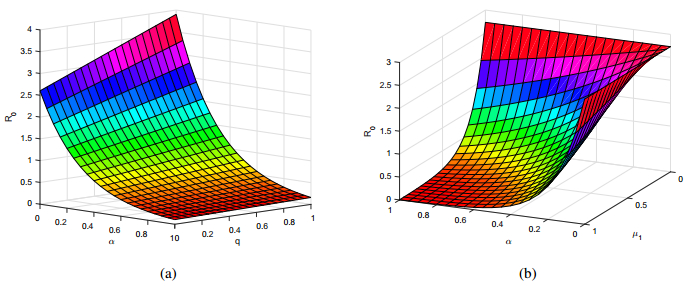
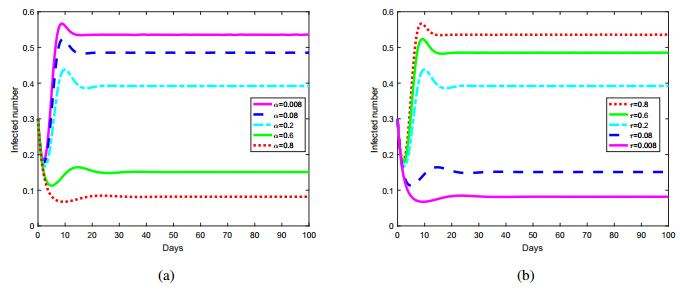
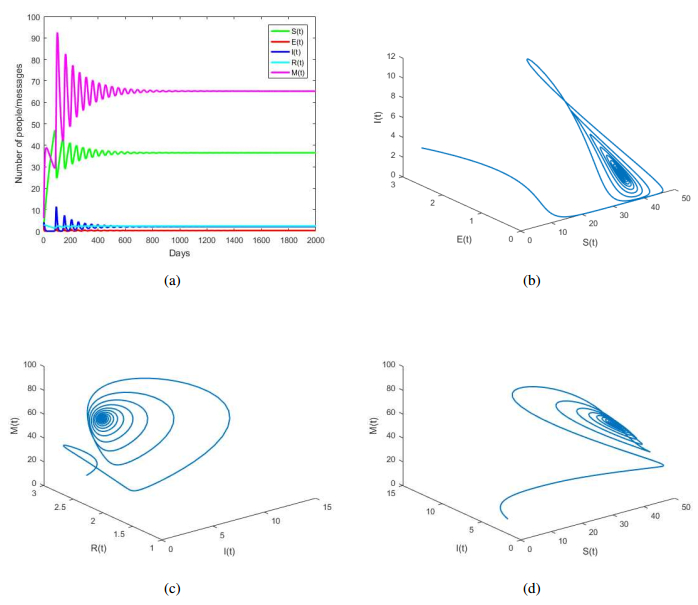
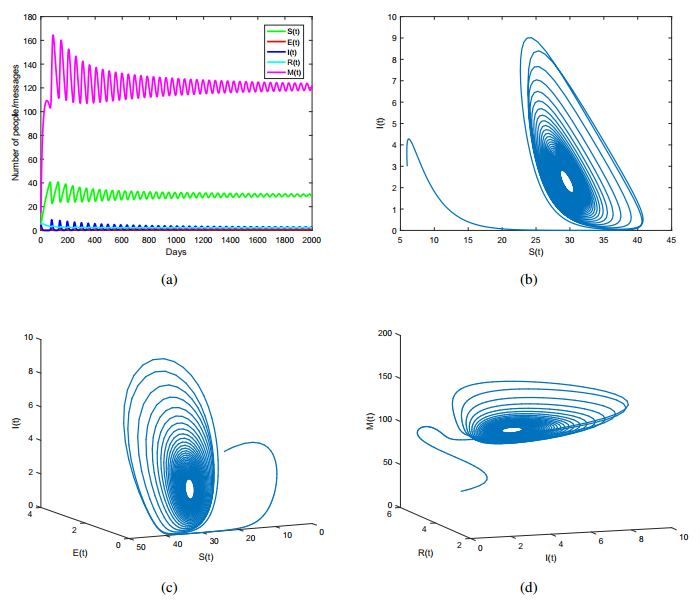
Figures(8) / Tables(1)

Ya-Dong Zhang, Hai-Feng Huo, Hong Xiang. Dynamics of tuberculosis with fast and slow progression and media coverage[J]. Mathematical Biosciences and Engineering, 2019, 16(3): 1150-1170. doi: 10.3934/mbe.2019055







 DownLoad:
DownLoad: