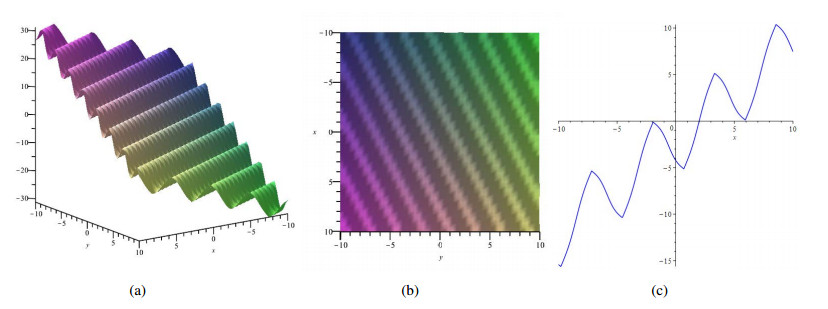
In this paper, a (2+1)-dimensional KdV4 equation is considered. We obtain Lie symmetries of this equation by utilizing Lie point symmetry analysis method, then use them to perform symmetry reductions. By using translation symmetries, two fourth-order ordinary differential equations are obtained. Solutions of one fourth order ordinary differential equation are presented by using direct integration method and $ (G'/G) $-expansion method respectively. Furthermore, the corresponding solutions are depicted with appropriate graphical representations. The other fourth-order ordinary differential equation is solved by using power series technique. Finally, two kinds of conserved vectors of this equation are presented by invoking the multiplier method and Noether's theorem respectively.
Citation: Sixing Tao. Lie symmetry analysis, particular solutions and conservation laws of a (2+1)-dimensional KdV4 equation[J]. Mathematical Biosciences and Engineering, 2023, 20(7): 11978-11997. doi: 10.3934/mbe.2023532
In this paper, a (2+1)-dimensional KdV4 equation is considered. We obtain Lie symmetries of this equation by utilizing Lie point symmetry analysis method, then use them to perform symmetry reductions. By using translation symmetries, two fourth-order ordinary differential equations are obtained. Solutions of one fourth order ordinary differential equation are presented by using direct integration method and $ (G'/G) $-expansion method respectively. Furthermore, the corresponding solutions are depicted with appropriate graphical representations. The other fourth-order ordinary differential equation is solved by using power series technique. Finally, two kinds of conserved vectors of this equation are presented by invoking the multiplier method and Noether's theorem respectively.

| [1] |
C. M. Khalique, K. Maefo, A study on the (2+1)-dimensional first extended Calogero-Bogoyavlenskii-Schiff equation, Math. Biosci. Eng., 18 (2021), 5816–5835. https://doi.org/10.3934/mbe.2021293 doi: 10.3934/mbe.2021293

|
| [2] |
S. F. Tian, Lie symmetry analysis, conservation laws and solitary wave solutions to a fourth-order nonlinear generalized Boussinesq water wave equation, Appl. Math. Lett., 100 (2020), 106056. https://doi.org/10.1016/j.aml.2019.106056 doi: 10.1016/j.aml.2019.106056
|
| [3] |
C. M. Khalique, O. D. Adeyemo, K. Maefo, Symmetry solutions and conservation laws of a new generalized 2D Bogoyavlensky-Konopelchenko equation of plasma physics, AIMS Math., 7 (2022), 9767–9788. https://doi.org/10.3934/math.2022544 doi: 10.3934/math.2022544
|
| [4] |
L. D. Moleleki, I. Simbanefayi, C. M. Khalique, Symmetry solutions and conservation laws of a (3+1)-dimensional generalized KP-Boussinesq equation in fluid mechanics, Chin. J. Phys., 68 (2020), 940–949. https://doi.org/10.1016/j.cjph.2020.10.023 doi: 10.1016/j.cjph.2020.10.023
|
| [5] |
W. X. Ma, Y. J. Zhang, Darboux transformations of integrable couplings and applications, Rev. Math. Phys., 30 (2018), 1850003. https://doi.org/10.1142/S0129055X18500034 doi: 10.1142/S0129055X18500034
|
| [6] |
D. Bilman, P. D. Miller, A robust inverse scattering transform for the focusing nonlinear Schrödinger equation, Commun. Pure Appl. Math., 72 (2019), 1722–1805. https://doi.org/10.1002/cpa.21819 doi: 10.1002/cpa.21819
|
| [7] |
S. K. Liu, Z. T. Fu, S. D. Liu, Q. Zhao, Jacobi elliptic function expansion method and periodic wave solutions of nonlinear wave equations, Phys. Lett. A, 289 (2001), 69–74. https://doi.org/10.1016/S0375-9601(01)00580-1 doi: 10.1016/S0375-9601(01)00580-1
|
| [8] |
N. A. Kudryashov, One method for finding exact solutions of nonlinear differential equations, Commun. Nonlinear Sci., 17 (2012), 2248–2253. https://doi.org/10.1016/j.cnsns.2011.10.016 doi: 10.1016/j.cnsns.2011.10.016
|
| [9] |
M. L. Wang, X. Z. Li, J. L. Zhang, The $(G^{'}/G)$-expansion method and travelling wave solutions of nonlinear evolution equations in mathematical physics, Phys. Lett. A, 372 (2008), 417–423. https://doi.org/10.1016/j.physleta.2007.07.051 doi: 10.1016/j.physleta.2007.07.051
|
| [10] |
Y. L. Zhao, Y. P. Liu, Z. B. Li, A connection between the $(G'/G)$-expansion method and the truncated Painleve expansion method and its application to the mKdV equation, Chin. Phys. B, 19 (2010), 030306. https://doi.org/10.1088/1674-1056/19/3/030306 doi: 10.1088/1674-1056/19/3/030306
|
| [11] |
S. Sirisubtawee, S. Koonprasert, Exact traveling wave solutions of certain nonlinear partial differential equations using the $(G'/G^{2})$-expansion method, Adv. Math. Phys., 2018 (2018), 7628651. https://doi.org/10.1155/2018/7628651 doi: 10.1155/2018/7628651
|
| [12] |
S. X. Tao, Nonlocal symmetry, CRE solvability, and exact interaction solutions of the (2+1)-dimensional Boussinesq equation, J. Math., 2022 (2022), 7850824. https://doi.org/10.1155/2022/7850824 doi: 10.1155/2022/7850824
|
| [13] |
C. M. Khalique, L. D. Moleleki, A (3+1)-dimensional generalized BKP-Boussinesq equation: Lie group approach, Results Phys., 13 (2019), 102239. https://doi.org/10.1016/j.rinp.2019.102239 doi: 10.1016/j.rinp.2019.102239
|
| [14] |
I. Simbanefayi, C. M. Khalique, Travelling wave solutions and conservation laws for the Korteweg-de Vries-Bejamin-Bona-Mahony equation, Results Phys., 8 (2018), 57–63. https://doi.org/10.1016/j.rinp.2017.10.041 doi: 10.1016/j.rinp.2017.10.041
|
| [15] |
H. Z. Liu, L. J. Zhang, Symmetry reductions and exact solutions to the systems of nonlinear partial differential equations, Phys. Scripta, 94 (2019), 015202. https://doi.org/10.1088/1402-4896/aaeeff doi: 10.1088/1402-4896/aaeeff
|
| [16] | G. W. Bluman, S. Kumei, Symmetries and Differential Equations, Springer-Verlag, New York, 1989. https://doi.org/10.2307/3619872 |
| [17] |
L. Ju, J. Zhou, Y. F. Zhang, Conservation laws analysis of nonlinear partial differential equations and their linear soliton solutions and Hamiltonian structures, Commun. Anal. Mech., 15 (2023), 24–49. https://doi.org/10.3934/cam.2023002 doi: 10.3934/cam.2023002
|
| [18] |
R. Naz, F. M. Mahomed, D. P. Mason, Comparison of different approaches to conservation laws for some partial differential equations in fluid mechanics, Appl. Math. Comput., 205 (2008), 212–230. https://doi.org/10.1016/j.amc.2008.06.042 doi: 10.1016/j.amc.2008.06.042
|
| [19] |
A. Sjöberg, On double reductions from symmetries and conservation laws, Nonlinear Anal. Real World Appl., 10 (2009), 3472–3477. https://doi.org/10.1016/j.nonrwa.2008.09.029 doi: 10.1016/j.nonrwa.2008.09.029
|
| [20] |
E. Noether, Invariant variation problems, Transp. Theory Stat. Phys., 1 (1971), 186–207. https://doi.org/10.1080/00411457108231446 doi: 10.1080/00411457108231446
|
| [21] |
W. Sarlet, Comment on 'Conservation laws of higher order nonlinear PDEs and the variational conservation laws in the class with mixed derivatives', J. Phys. A: Math. Theor., 43 (2010), 458001. https://doi.org/10.1088/1751-8113/43/45/458001 doi: 10.1088/1751-8113/43/45/458001
|
| [22] |
T. Motsepa, M. Abudiab, C. M. Khalique, A study of an extended generalized (2+1)-dimensional Jaulent-Miodek equation, Int. J. Nonlinear Sci. Numer. Simul., 19 (2018), 391–395. https://doi.org/10.1515/ijnsns-2017-0147 doi: 10.1515/ijnsns-2017-0147
|
| [23] |
N. H. Ibragimov, A new conservation theorem, J. Math. Anal. Appl., 333 (2007), 311–328. https://doi.org/10.1016/j.jmaa.2006.10.078 doi: 10.1016/j.jmaa.2006.10.078
|
| [24] |
C. M. Khalique, S. A. Abdallah, Coupled Burgers equations governing polydispersive sedimentation; a Lie symmetry approach, Results Phys., 16 (2020), 102967. https://doi.org/10.1016/j.rinp.2020.102967 doi: 10.1016/j.rinp.2020.102967
|
| [25] |
M. S. Bruzón, M. L. Gandarias, Traveling wave solutions of the $K(m, n)$ equation with generalized evolution, Math. Meth. Appl. Sci., 41 (2018), 5851–5857. https://doi.org/10.1002/mma.1339 doi: 10.1002/mma.1339
|
| [26] | P. J. Olver, Applications of Lie Groups to Differential Equations, 2$^nd$ edition, Springer-Verlag, New York, 1993. https://doi.org/10.1007/978-1-4684-0274-2 |
| [27] |
X. Li, L. Wang, Z. J. Zhou, Y. Chen, Z. Y. Yan, Stable dynamics and excitations of single- and double-hump solitons in the Kerr nonlinear media with $\mathcal PT$-symmetric HHG potentials, Nonlinear Dyn., 108 (2022), 4045–4056. https://doi.org/10.1007/s11071-022-07362-1 doi: 10.1007/s11071-022-07362-1
|
| [28] |
G. Q. Xu, A. M. Wazwaz, Bidirectional solitons and interaction solutions for a new integrable fifth-order nonlinear equation with temporal and spatial dispersion, Nonlinear Dyn., 101 (2020), 581–595. https://doi.org/10.1007/s11071-020-05740-1 doi: 10.1007/s11071-020-05740-1
|
| [29] |
J. G. Liu, X. J. Yang, Y. Y. Feng, P. Cui, Nonlinear dynamic behaviors of the generalized (3+1)-dimensional KP equation, ZAMM-J. Appl. Math. Mech., 102 (2022), e202000168. https://doi.org/10.1002/zamm.202000168 doi: 10.1002/zamm.202000168
|
| [30] |
A. M. Wazwaz, A study on the (2+1)-dimensional KdV4 equation derived by using the KdV recursion operator, Math. Methods Appl. Sci., 36 (2013), 1760–1767. https://doi.org/10.1002/mma.2721 doi: 10.1002/mma.2721
|
| [31] |
Y. P. Wang, B. Tian, M. Wang, Y. F. Wang, Y. Sun, X. Y. Xie, Bäcklund transformations and soliton solutions for a (2+1)-dimensional Korteweg–de Vries-type equation in water waves, Nonlinear Dyn., 81 (2015), 1815–1821. https://doi.org/10.1007/s11071-015-2109-y doi: 10.1007/s11071-015-2109-y
|
| [32] |
S. X. Tao, Breather wave and traveling wave solutions for a (2+1)-dimensional KdV4 equation, Adv. Math. Phys., 2022 (2022), 7761659. https://doi.org/10.1155/2022/7761659 doi: 10.1155/2022/7761659
|





Figures(6)

Sixing Tao. Lie symmetry analysis, particular solutions and conservation laws of a (2+1)-dimensional KdV4 equation[J]. Mathematical Biosciences and Engineering, 2023, 20(7): 11978-11997. doi: 10.3934/mbe.2023532







 DownLoad:
DownLoad: