Citation: Manel Dali Youcef, Alain Rapaport, Tewfik Sari. Study of performance criteria of serial configuration of two chemostats[J]. Mathematical Biosciences and Engineering, 2020, 17(6): 6278-6309. doi: 10.3934/mbe.2020332

| [1] | J. Monod, La technique de culture continue: Theorie et applications, Ann. Inst. Pasteur Mic., 79 (1950), 39-410. |
| [2] | A. Novick, L. Szilard, Description of the chemostat, Science, 112 (1950), 715-716. |
| [3] | D. Herbert, R. Elsworth, R. C. Telling, The continuous culture of bacteria; a theoretical and experimental study, Microbiology, 14 (1956), 601-622. |
| [4] | J. Harmand, C. Lobry, A. Rapaport, T. Sari, The Chemostat: Mathematical Theory of Microorganism Cultures, John Wiley & Sons, 2017. |
| [5] |
P. A. Hoskisson, G. Hobbs, Continuous culture-making a comeback?, Microbiology, 151 (2005), 3153-3159. doi: 10.1099/mic.0.27924-0

|
| [6] | H. L. Smith, P. Waltman, The theory of the chemostat: dynamics of microbial competition, Cambridge University Press, Cambridge, 1995. |
| [7] | M. Wade, J. Harmand, B. Benyahia, T. Bouchez, S. Chaillou, B. Cloez, et al., Perspectives in mathematical modelling for microbial ecology, Ecol. Model., 321 (2016), 64-74. |
| [8] | C. P. L. Grady, G. T. Daigger, N. G. Love, C. D. M. Filipe, Biological wastewater treatment, 3rd edition, CRC press, 2011. |
| [9] | D. Dochain, P. A. Vanrolleghem, Dynamic modelling & estimation in wastewater treatment processes, IWA Publishing, (2001). |
| [10] |
C. M. Kung, B. Baltzis, The growth of pure and simple microbial competitors in a moving and distributed medium, Math. Biosci., 111 (1992), 295-313. doi: 10.1016/0025-5564(92)90076-9
|
| [11] |
B. Tang, Mathematical investigations of growth of microorganisms in the gradostat, J. Math. Biol., 23 (1986), 319-339. doi: 10.1007/BF00275252
|
| [12] |
C. D. de Gooijer, W. A. M. Bakker Wilfried, H. H. Beeftink, J. Tramper, Bioreactors in series: An overview of design procedures and practical applications, Enzyme Microb. Tech., 18 (1996), 202-219. doi: 10.1016/0141-0229(95)00090-9
|
| [13] |
G. Hill, C. Robinson, Minimum tank volumes for CFST bioreactors in series, Can. J. Chem. Eng., 67 (1989), 818-824. doi: 10.1002/cjce.5450670513
|
| [14] |
E. Scuras, A. Jobbagy, L. Grady, Optimization of activated sludge reactor configuration: kinetic considerations, Water Res., 35 (2001), 4277-4284. doi: 10.1016/S0043-1354(01)00177-4
|
| [15] |
A. Rapaport, Some non-intuitive properties of simple extensions of the chemostat model, Ecol. Complexity, Elsevier, 34 (2018), 111-118. doi: 10.1016/j.ecocom.2017.02.003
|
| [16] |
I. Haidar, A. Rapaport, F. Gérard, Effects of spatial structure and diffusion on the performances of the chemostat, Math. Biosci. Eng., 8 (2011), 953-971. doi: 10.3934/mbe.2011.8.953
|
| [17] | J. Harmand, Contribution à l'analyse et au contrôle des systèmes biologiques application aux bio-procédés de dépollution, Habilitation à diriger des recherches, Université C. Bernard Lyon, 2004. |
| [18] | J. Zambrano, B. Carlsson, Optimizing zone volumes in bioreactors described by Monod and Contois growth kinetics, Proceeding of the IWA World Water Congress & Exhibition, 2014. Available from: https://www.researchgate.net/profile/Jesus_Zambrano4/publication/261831366. |
| [19] |
J. Zambrano, B. Carlsson, S. Diehl, Optimal steady-state design of zone volumes of bioreactors with Monod growth kinetics, Biochem. Eng. J., 100 (2015), 59-66. doi: 10.1016/j.bej.2015.04.002
|
| [20] | D. Herbert, Multi-stage continuous culture, in Continuous cultivation of microorganism, Academic Press, 1964. |
| [21] | I. Haidar, Dynamiques Microbiennes Et Modélisation Des Cycles Biogéochimiques Terrestres, Thèse de l'Université de Montpellier, 2011. |
| [22] |
R. Fekih-Salem, C. Lobry, T. Sari, A density-dependent model of competition for one resource in the chemostat, Math. Biosci., 286 (2017), 104-122. doi: 10.1016/j.mbs.2017.02.007
|
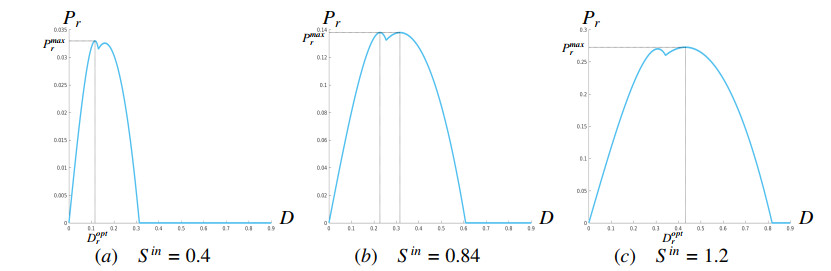
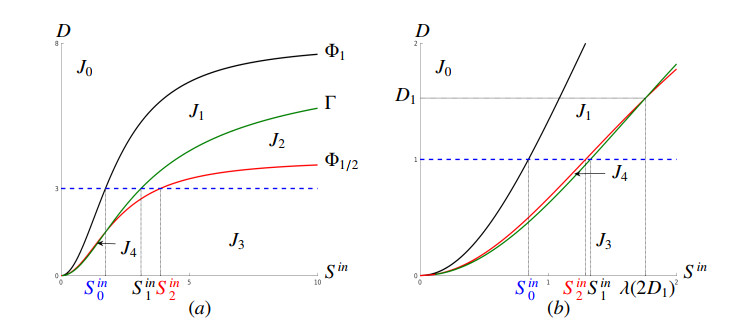
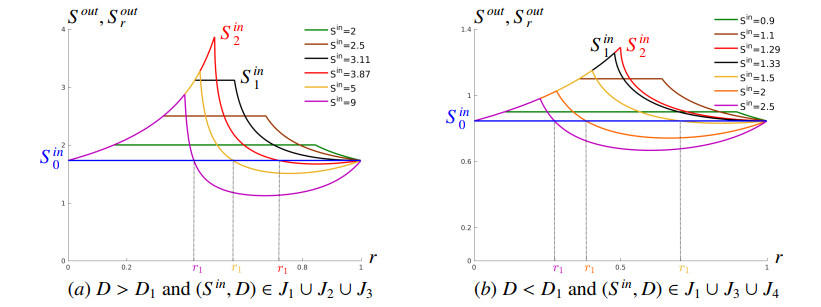
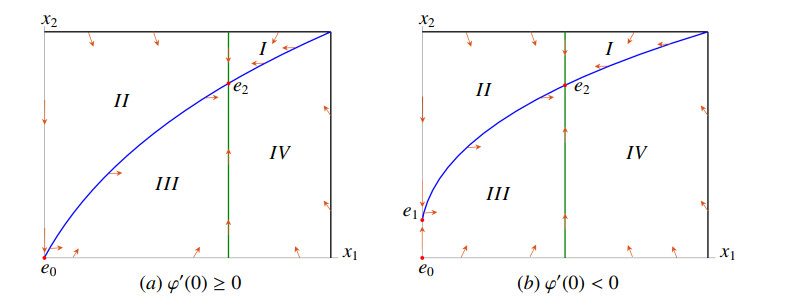
Figures(16) / Tables(2)

Manel Dali Youcef, Alain Rapaport, Tewfik Sari. Study of performance criteria of serial configuration of two chemostats[J]. Mathematical Biosciences and Engineering, 2020, 17(6): 6278-6309. doi: 10.3934/mbe.2020332







 DownLoad:
DownLoad: