|
[1]
|
GBD 2013 Mortality and Causes of Death Collaborators, Global, regional, and national age–sex specific all-cause and cause-specific mortality for 240 causes of death, 1990–2013: A systematic analysis for the Global Burden of Disease Study 2013, Lancet., 385 (2015), 117–171.
|
|
[2]
|
B.K. Yoo, L. Emdad, Z. Su, et al., Astrocyte elevated gene-1 regulates hepatocellular carcinoma development and progression, J. Clin. Invest., 119 (2009), 465–477.
|
|
[3]
|
N. Portolani, A. Coniglio, S. Ghidoni, et al., Early and late recurrence after liver resection for hepatocellular carcinoma: Prognostic and therapeutic implications., Ann. Surg., 243 (2006), 229–235.
|
|
[4]
|
C.E. Brinckerhoff and L.M. Matrisian, TIMELINEMatrix metalloproteinases: A tail of a frog that became a prince, Nat. Rev. Mol. Cell Biol., 3 (2002), 207–214.
|
|
[5]
|
R. Chen, J. Cui, C. Xu, et al., The Significance of MMP-9 Over MMP-2 in HCC Invasiveness and Recurrence of Hepatocellular Carcinoma After Curative Resection, Ann. Surg. Oncol., 19 (2012), 375–384.
|
|
[6]
|
C. Gialeli, A.D. Theocharis and N.K. Karamanos, Roles of matrix metalloproteinases in cancer progression and their pharmacological targeting, FEBS J., 278 (2011), 16–27.
|
|
[7]
|
E.I. Deryugina and J.P. Quigley, Matrix metalloproteinases and tumor metastasis, Cancer Metastasis Rev., 25 (2006), 9–34.
|
|
[8]
|
R. Derynck, R.J. Akhurst and A. Balmain, TGF-|[beta]| signaling in tumor suppression and cancer progression, Nat. Genet., 29 (2001), 117–129.
|
|
[9]
|
Y. Katsuno, S. Lamouille and R. Derynck, TGF-β signaling and epithelial–mesenchymal transition in cancer progression, Curr. Opin. Oncol., 25 (2013), 76–84.
|
|
[10]
|
A. Safina, M.Q. Ren, E. Vandette, et al., TAK1 is required for TGF-β1-mediated regulation of matrix metalloproteinase-9 and metastasis, Oncogene., 27 (2008), 1198–1207.
|
|
[11]
|
S.L. Dallas, J.L. Rosser, G.R. Mundy, et al., Proteolysis of latent transforming growth factor-beta (TGF-beta )-binding protein-1 by osteoclasts. A cellular mechanism for release of TGF-beta from bone matrix, J. Biol. Chem., 277 (2002), 21352–21360.
|
|
[12]
|
J. Li, B. Yang, Q. Zhou, et al., Autophagy promotes hepatocellular carcinoma cell invasion through activation of epithelial–mesenchymal transition, Carcinogenesis, 34 (2013), 1343–1351.
|
|
[13]
|
G. Chen, G. Qin, Y. Dang, et al., The prospective role of matrix metalloproteinase-2/9 and transforming growth factor beta 1 in accelerating the progression of hepatocellular carcinoma, Transl. Cancer Res., 6 (2017), S229–S231.
|
|
[14]
|
A. V. Bakin, A.K. Tomlinson, N.A. Bhowmick, et al., Phosphatidylinositol 3-Kinase Function Is Required for Transforming Growth Factor β-mediated Epithelial to Mesenchymal Transition and Cell Migration, J. Biol. Chem., 275 (2000), 36803–36810.
|
|
[15]
|
J.H. Zuo, W. Zhu, M.Y. Li, et al., Activation of EGFR promotes squamous carcinoma SCC10A cell migration and invasion via inducing EMT-like phenotype change and MMP-9-mediated degradation of E-cadherin, J. Cell. Biochem., 112 (2011), 2508–2517.
|
|
[16]
|
J. Krstic and J.F. Santibanez, Transforming Growth Factor-Beta and Matrix Metalloproteinases Functional Interplay in Cancer; Implications in Epithelial to Mesenchymal Transition, Cell Biol. Res. Ther., s1 (2016).
|
|
[17]
|
R. Bataller and D.A. Brenner, Liver fibrosis, J. Clin. Invest., 115 (2005), 209–218.
|
|
[18]
|
H. de Jong, Modeling and Simulation of Genetic Regulatory Systems: A Literature Review, J. Comput. Biol., 9 (2002), 67–103.
|
|
[19]
|
L. Glass and S.A. Kauffman, The logical analysis of continuous, non-linear biochemical control networks., J. Theor. Biol., 39 (1973), 103–29.
|
|
[20]
|
H. DEJONG, J. GOUZE, C. HERNANDEZ, et al., Qualitative simulation of genetic regulatory networks using piecewise-linear models, Bull. Math. Biol., 66 (2004), 301–340.
|
|
[21]
|
R. Thomas and R. D'Ari, Biological feedback, CRC Press, 1990. https://www.crcpress.com/Biological-Feedback/Thomas-DAri/p/book/9780849367663.
|
|
[22]
|
E.M. Clarke, O. Grumberg and D.A. Peled, Model checking, MIT Press, 1999. https://mitpress.mit.edu/books/model-checking.
|
|
[23]
|
D.B. Z. Khalis, J. P. Comet and A. Richard, The SMBioNet Method for Discovering Models of Gene Regulatory Networks, Genes, Genomes and Genomics., 1 (2009), 15–22.
|
|
[24]
|
G. Bernot, J.P. Comet, A. Richard, et al., Application of formal methods to biological regulatory networks: extending Thomas' asynchronous logical approach with temporal logic, J. Theor. Biol., 229 (2004), 339–34 7.
|
|
[25]
|
A. Cimatti, E.M. Clarke E. Giunchiglia, et al., NuSMV 2: An OpenSource Tool for Symbolic Model Checking, (n.d.). http://repository.cmu.edu/compsci.
|
|
[26]
|
A.G. Gonzalez, A. Naldi, L. Sánchez, et al., GINsim: A software suite for the qualitative modelling, simulation and analysis of regulatory networks, Biosystems, 84 (2006), 91–100.
|
|
[27]
|
J. Ahmad, Modélisation hybride et analyse des dynamiques des réseaux de régulations biologiques en tenant compte des délais, PhD Thesis, Nantes., (2009).
|
|
[28]
|
M. Heiner, M. Herajy, F. Liu, et al., Snoopy – A Unifying Petri Net Tool, (2012), 398–407.
|
|
[29]
|
A. Naldi, D. Berenguier, A. Fauré, et al., Logical modelling of regulatory networks with GINsim 2.3, Biosystems, 97 (2009), 134–139.
|
|
[30]
|
A. Saadatpour, R. Albert and T.C. Reluga, A Reduction Method for Boolean Network Models Proven to Conserve Attractors, SIAM J. Appl. Dyn. Syst., 12 (2013), 1997–2011.
|
|
[31]
|
E. Plahte, T. Mestl and S.W. Omholt, Feedback loops, stability and multistationarity in dynamical systems, J. Biol. Syst., 03 (1995), 409–413.
|
|
[32]
|
D. Bissell, D. Roulot and J. George, Transforming growth factor β and the liver, Hepatology, 34 (2001), 859–867.
|
|
[33]
|
G. Chen, G. Qin, Y. Dang, et al., The prospective role of matrix metalloproteinase-2/9 and transforming growth factor beta 1 in accelerating the progression of hepatocellular carcinoma, Transl. Cancer Res., 6 (2017), S229–S231.
|
|
[34]
|
M. Bond, A.J. Chase, A.H. Baker, et al., Inhibition of transcription factor NF-kappaB reduces matrix metalloproteinase-1, -3 and -9 production by vascular smooth muscle cells., Cardiovasc. Res., 50 (2001), 556–565.
|
|
[35]
|
H. Nakatsukasa, Cellular distribution of transcripts for tissue inhibitor of metalloproteinases 1 and 2 in human hepatocellular carcinomas, Hepatology, 24 (1996), 82–88.
|
|
[36]
|
P. Shannon, A. Markiel, O. Ozier, et al., Cytoscape: a software environment for integrated models of biomolecular interaction networks., Genome Res., 13 (2003), 2498–504.
|
|
[37]
|
A. Keutgens, I. Robert, P. Viatour, et al., Deregulated NF-kappaB activity in haematological malignancies., Biochem. Pharmacol., 72 (2006), 1069–80.
|
|
[38]
|
W.G. Stetler-Stevenson, Matrix metalloproteinases in angiogenesis: A moving target for therapeutic intervention., J. Clin. Invest., 103 (1999), 1237–1241.
|
|
[39]
|
A. Ma'ayan, Introduction to network analysis in systems biology, Sci. Signal., 4 (2011), tr5.
|
|
[40]
|
O. Palmieri, T. Mazza, S. Castellana, et al., Inflammatory Bowel Disease Meets Systems Biology: A Multi-Omics Challenge and Frontier, Omi. A J. Integr. Biol., 20 (2016), 692–698.
|
|
[41]
|
G. Giannelli, E. Villa and M. Lahn, Transforming growth factor-β as a therapeutic target in hepatocellular carcinoma, Cancer Res., 74 (2014), 1890–1894.
|
|
[42]
|
G. Giannelli, A. Mazzocca, E. Fransvea, et al., Inhibiting TGF-β signaling in hepatocellular carcinoma, Biochim. Biophys. Acta - Rev. Cancer., 1815 (2011), 214–223.
|
|
[43]
|
J. Wang, C.-P. Zhu, P.-F. Hu, et al., FOXA2 suppresses the metastasis of hepatocellular carcinoma partially through matrix metalloproteinase-9 inhibition, Carcinogenesis, 35 (2014), 2576–2583.
|
|
[44]
|
H.B. Yu, H.F. Zhang, D.Y. Li, et al., Journal of Asian Natural Products Research Matrine inhibits matrix metalloproteinase-9 expression and invasion of human hepatocellular carcinoma cells Matrine inhibits matrix metalloproteinase-9 expression and invasion of human hepatocellular carcinoma cells, J. Asian Nat. Prod. Res., 13 (2011), 3–242.
|
|
[45]
|
C.F. Huang, Y.H. Teng, F.J. Lu, et al., β-mangostin suppresses human hepatocellular carcinoma cell invasion through inhibition of MMP-2 and MMP-9 expression and activating the ERK and JNK pathways, Environ. Toxicol., 32 (2017), 2360–2370.
|
|
[46]
|
M. Egeblad and Z. Werb, New functions for the matrix metalloproteinases in cancer progression, Nat. Rev. Cancer., 2 (2002), 161–174.
|
|
[47]
|
M.D. Sternlicht and Z. Werb, How Matrix Metalloproteinases Regulate Cell Behavior, Annu. Rev. Cell Dev. Biol., 17 (2001), 463–516.
|
|
[48]
|
G. Qin, M. Luo, J. Chen, et al., Reciprocal activation between MMP-8 and TGF-β1 stimulates EMT and malignant progression of hepatocellular carcinoma, Cancer Lett., 374 (2016), 85–95.
|
|
[49]
|
H. Kitano, Looking beyond the details: A rise in system-oriented approaches in genetics and molecular biology, Curr. Genet., 41 (2002), 1–10.
|
|
[50]
|
D. THIEFFRY and R. THOMAS, Dynamical behaviour of biological regulatory networks-II. Immunity control in bacteriophage lambda*, Bull. Math. Biol., 57 (1995), 277–297.
|
|
[51]
|
V.K. Rakyan, T.A. Down, D.J. Balding, et al., Epigenome-wide association studies for common human diseases, Nat. Rev. Genet., 12 (2011), 529–541.
|
|
[52]
|
B. BIERIE and H. MOSES, TGF-β and cancer, Cytokine Growth Factor Rev., 17 (2006), 29–40.
|
|
[53]
|
A. Noël, M. Jost and E. Maquoi, Matrix metalloproteinases at cancer tumor–host interface, Semin. Cell Dev. Biol., 19 (2008), 52–60.
|
|
[54]
|
K. Kessenbrock, V. Plaks and Z. Werb, Matrix Metalloproteinases: Regulators of the Tumor Microenvironment, Cell, 141 (2010), 52–67.
|

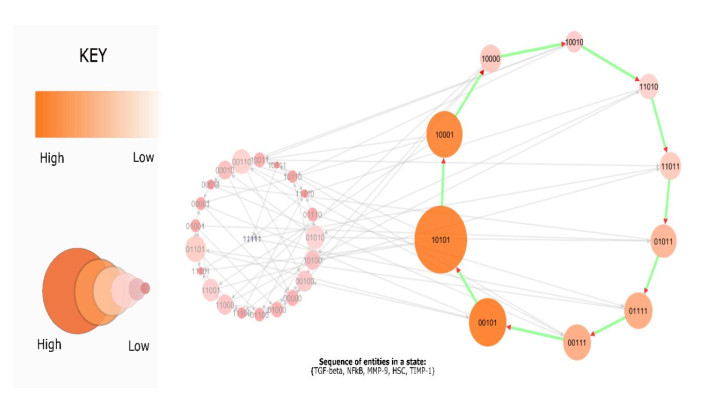
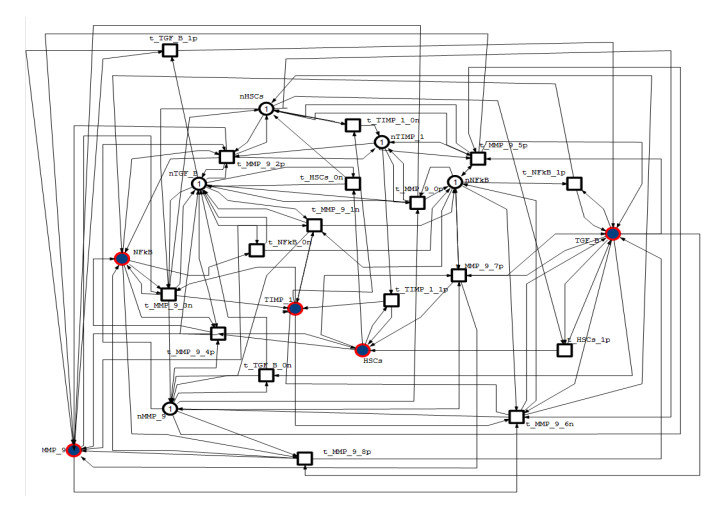
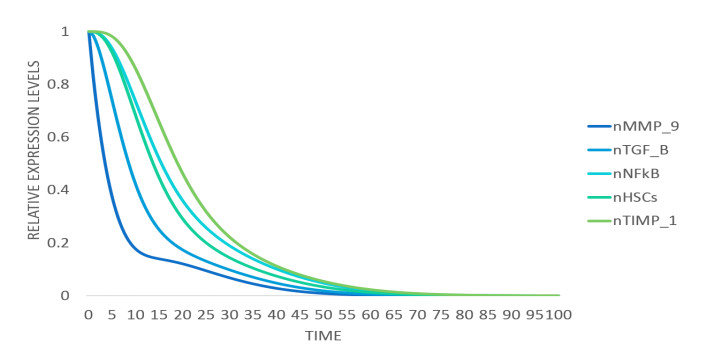
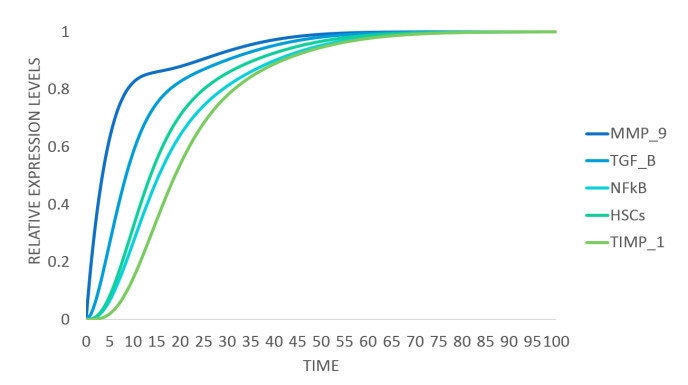








 DownLoad:
DownLoad: