We aimed to explore the expression and clinical prognostic significance of PAQR4 in hepatocellular carcinoma (HCC).
We obtained the gene expression matrix and clinical data of HCC from the cancer genome atlas (TCGA) and international cancer genome consortium (ICGC) databases. The prognostic value of PAQR4 in HCC was evaluated using the Kaplan-Meier and Cox regression analyses. PAQR4-related pathways were explored by gene set enrichment analysis (GSEA). A clinical nomogram prognostic model based on the PAQR family was constructed using Cox proportional hazards models.
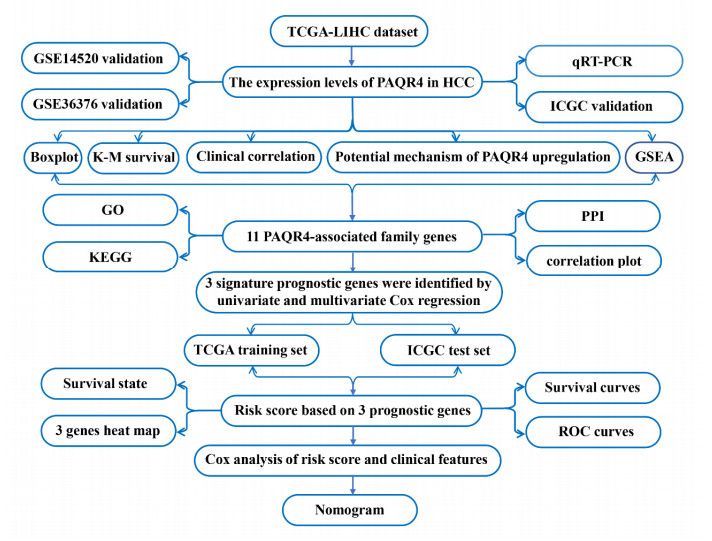
We found that PAQR4 is overexpressed in HCC from multiple databases; additionally, quantitative real-time polymerase chain reaction (qRT-PCR) validated the upregulation of PAQR4 in HCC. PAQR4 expression was related to age, grade, alpha fetoprotein (AFP), T classification and clinical stage of HCC patients. High PAQR4 expression was associated with poor overall survival and was an independent prognostic factor for HCC patients through Kaplan-Meier analysis and Cox regression analysis, respectively. In addition, GSEA identified that the high PAQR4 expression phenotype was involved in the cell cycle, Notch signaling pathway, mTOR signaling pathway, etc. Finally, three PAQR family genes (PAQR4, PAQR8 and PAQR9) were associated with the prognosis of patients with HCC. A clinical nomogram prediction model was verified in TCGA training and ICGC validation sets, and it exerted dramatic predictive efficiency in this study.
PAQR4 may be regarded as a promising prognostic biomarker and therapeutic target for HCC.
Citation: Caihao Qu, Tengda Ma, Xin YAN, Xiaomei Li, Yumin Li. Overexpressed PAQR4 predicts poor overall survival and construction of a prognostic nomogram based on PAQR family for hepatocellular carcinoma[J]. Mathematical Biosciences and Engineering, 2022, 19(3): 3069-3090. doi: 10.3934/mbe.2022142
We aimed to explore the expression and clinical prognostic significance of PAQR4 in hepatocellular carcinoma (HCC).
We obtained the gene expression matrix and clinical data of HCC from the cancer genome atlas (TCGA) and international cancer genome consortium (ICGC) databases. The prognostic value of PAQR4 in HCC was evaluated using the Kaplan-Meier and Cox regression analyses. PAQR4-related pathways were explored by gene set enrichment analysis (GSEA). A clinical nomogram prognostic model based on the PAQR family was constructed using Cox proportional hazards models.
We found that PAQR4 is overexpressed in HCC from multiple databases; additionally, quantitative real-time polymerase chain reaction (qRT-PCR) validated the upregulation of PAQR4 in HCC. PAQR4 expression was related to age, grade, alpha fetoprotein (AFP), T classification and clinical stage of HCC patients. High PAQR4 expression was associated with poor overall survival and was an independent prognostic factor for HCC patients through Kaplan-Meier analysis and Cox regression analysis, respectively. In addition, GSEA identified that the high PAQR4 expression phenotype was involved in the cell cycle, Notch signaling pathway, mTOR signaling pathway, etc. Finally, three PAQR family genes (PAQR4, PAQR8 and PAQR9) were associated with the prognosis of patients with HCC. A clinical nomogram prediction model was verified in TCGA training and ICGC validation sets, and it exerted dramatic predictive efficiency in this study.
PAQR4 may be regarded as a promising prognostic biomarker and therapeutic target for HCC.

| [1] |
H. Sung, J. Ferlay, R. L. Siegel, M. Laversanne, I. Soerjomataram, A. Jemal, et al., Global cancer statistics 2020: globocan estimates of incidence and mortality worldwide for 36 cancers in 185 countries, CA Cancer J. Clin., 71 (2021), 209–249. https://doi.org/10.3322/caac.21660 doi: 10.3322/caac.21660

|
| [2] |
F. Bray, J. Ferlay, I. Soerjomataram, R. L. Siegel, L. A. Torre, A. Jemal, Global cancer statistics 2018: globocan estimates of incidence and mortality worldwide for 36 cancers in 185 countries, CA Cancer J. Clin., 68 (2018), 394–424. https://doi.org/10.3322/caac.21492 doi: 10.3322/caac.21492
|
| [3] |
M. Xing, X. Wang, R. A. Kiken, L. He, J. Y. Zhang, Immunodiagnostic biomarkers for hepatocellular carcinoma (HCC): the first step in detection and treatment, Int. J. Mol. Sci., 22 (2021), 6139. https://doi.org/10.3390/ijms22116139 doi: 10.3390/ijms22116139
|
| [4] |
S. Temraz, F. Nassar, F. Kreidieh, D. Mukherji, A. Shamseddine, R. Nasr, Hepatocellular carcinoma immunotherapy and the potential influence of gut microbiome, Int. J. Mol. Sci., 22 (2021), 7800. https://doi.org/10.3390/ijms22157800 doi: 10.3390/ijms22157800
|
| [5] |
A. Rizzo, A. D. Ricci, PD-L1, TMB, and other potential predictors of response to immunotherapy for hepatocellular carcinoma: how can they assist drug clinical trials?, Expert Opin. Inv. Drug., 30 (2021), 1–9. https://doi.org/10.1080/13543784.2021.1972969 doi: 10.1080/13543784.2021.1972969
|
| [6] |
A. Rizzo, A. D. Ricci, G. Brandi, Atezolizumab in advanced hepatocellular carcinoma: good things come to those who wait, Immunotherapy, 13 (2021), 637–644. https://doi.org/10.2217/imt-2021-0026 doi: 10.2217/imt-2021-0026
|
| [7] |
A. Rizzo, V. Dadduzio, A. D. Ricci, F. Massari, A. Di Federico, G. Gadaleta-Caldarola, et al., Lenvatinib plus pembrolizumab: the next frontier for the treatment of hepatocellular carcinoma?, Expert Opin. Inv. Drug., 30 (2021), 1–8. https://doi.org/10.1080/13543784.2021.1948532 doi: 10.1080/13543784.2021.1948532
|
| [8] |
A. Rizzo, A. D. Ricci, G. Brandi, Immune-based combinations for advanced hepatocellular carcinoma: shaping the direction of first-line therapy, Future Oncol., 17 (2021), 755–757. https://doi.org/10.2217/fon-2020-0986 doi: 10.2217/fon-2020-0986
|
| [9] |
Y. T. Tang, T. Hu, M. Arterburn, B. Boyle, J. M. Bright, P. C. Emtage, et al., PAQR proteins: a novel membrane receptor family defined by an ancient 7-transmembrane pass motif, J. Mol. Evol., 61 (2005), 372–380. https://doi.org/10.1007/s00239-004-0375-2 doi: 10.1007/s00239-004-0375-2
|
| [10] |
P. Valadez-Cosmes, E. R. Vázquez-Martínez, M. Cerbón, I. Camacho-Arroyo, Membrane progesterone receptors in reproduction and cancer, Mol. Cell. Endocrinol., 434 (2016), 166–175. https://doi.org/10.1016/j.mce.2016.06.027 doi: 10.1016/j.mce.2016.06.027
|
| [11] |
L. Tian, N. Luo, X. Zhu, B. H. Chung, W. T. Garvey, Y. Fu, Adiponectin-AdipoR1/2-APPL1 signaling axis suppresses human foam cell formation: differential ability of AdipoR1 and AdipoR2 to regulate inflammatory cytokine responses, Atherosclerosis, 221 (2012), 66–75. https://doi.org/10.1016/j.atherosclerosis.2011.12.014 doi: 10.1016/j.atherosclerosis.2011.12.014
|
| [12] |
L. Lei, Z. N. Ling, X. L. Chen, L. L. Hong, Z. Q. Ling, Characterization of the Golgi scaffold protein PAQR3, and its role in tumor suppression and metabolic pathway compartmentalization, Cancer Manag. Res., 12 (2020), 353–362. https://doi.org/10.2147/CMAR.S210919 doi: 10.2147/CMAR.S210919
|
| [13] |
M. Sinreih, T. Knific, P. Thomas, S. Frković Grazio, T. L. Rižner, Membrane progesterone receptors β and γ have potential as prognostic biomarkers of endometrial cancer, J. Steroid Biochem. Mol. Biol., 178 (2018), 303–311. https://doi.org/10.1016/j.jsbmb.2018.01.011 doi: 10.1016/j.jsbmb.2018.01.011
|
| [14] |
T. Jin, Q. Ding, H. Huang, D. Xu, Y. Jiang, B. Zhou, et al., PAQR10 and PAQR11 mediate Ras signaling in the Golgi apparatus, Cell Res., 22 (2012), 661–676. https://doi.org/10.1038/cr.2011.161 doi: 10.1038/cr.2011.161
|
| [15] |
J. Ye, M. Gao, X. Guo, H. Zhang, F. Jiang, Breviscapine suppresses the growth and metastasis of prostate cancer through regulating PAQR4-mediated PI3K/Akt pathway, Biomed. Pharmacother., 127 (2020), 110223. https://doi.org/10.1016/j.biopha.2020.110223 doi: 10.1016/j.biopha.2020.110223
|
| [16] |
L. Pedersen, P. Panahandeh, M. I. Siraji, S. Knappskog, P. E. Lønning, R. Gordillo, et al., Golgi-localized PAQR4 mediates antiapoptotic ceramidase activity in breast cancer, Cancer Res., 80 (2020), 2163–2174. https://doi.org/10.1158/0008-5472.CAN-19-3177 doi: 10.1158/0008-5472.CAN-19-3177
|
| [17] |
P. Xu, L. Jiang, Y. Yang, M. Wu, B. Liu, Y. Shi, et al., PAQR4 promotes chemoresistance in non-small cell lung cancer through inhibiting Nrf2 protein degradation, Theranostics, 10 (2020), 3767–3778. https://doi.org/10.7150/thno.43142 doi: 10.7150/thno.43142
|
| [18] |
M. E. Ritchie, B. Phipson, D. Wu, Y. Hu, C.W. Law, W. Shi, et al., limma powers differential expression analyses for RNA-sequencing and microarray studies, Nucleic Acids Res., 43 (2015), e47. https://doi.org/10.1093/nar/gkv007 doi: 10.1093/nar/gkv007
|
| [19] |
E. Cerami, J. Gao, U. Dogrusoz, B. E. Gross, S. O. Sumer, B. A. Aksoy, et al., The cBio cancer genomics portal: an open platform for exploring multidimensional cancer genomics data, Cancer Discov., 2 (2012), 401–404. https://doi.org/10.1158/2159-8290.CD-12-0095 doi: 10.1158/2159-8290.CD-12-0095
|
| [20] |
C. Sticht, C. De La Torre, A. Parveen, N. Gretz, miRWalk: an online resource for prediction of microRNA binding sites, PLoS One, 13 (2018), e0206239. https://doi.org/10.1371/journal.pone.0206239 doi: 10.1371/journal.pone.0206239
|
| [21] |
V. Agarwal, G.W. Bell, J. W. Nam, D. P. Bartel, Predicting effective microRNA target sites in mammalian mRNAs, Elife, 4 (2015), e05005. https://doi.org/10.7554/eLife.05005 doi: 10.7554/eLife.05005
|
| [22] |
T. Li, J. Fan, B. Wang, N. Traugh, Q. Chen, J. S. Liu, et al., TIMER: a web server for comprehensive analysis of tumor-infiltrating immune cells, Cancer Res., 77 (2017), e108–e110. https://doi.org/10.1158/0008-5472.CAN-17-0307 doi: 10.1158/0008-5472.CAN-17-0307
|
| [23] |
A. Subramanian, P. Tamayo, V. K. Mootha, S. Mukherjee, B. L. Ebert, M. A. Gillette, et al., Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles, Proc. Natl. Acad. Sci. USA, 102 (2005), 15545–15550. https://doi.org/10.1073/pnas.0506580102 doi: 10.1073/pnas.0506580102
|
| [24] |
X. Wang, A. Spandidos, H. Wang, B. Seed, PrimerBank: a PCR primer database for quantitative gene expression analysis, 2012 update, Nucleic Acids Res., 40 (2012), D1144–D1149. https://doi.org/10.1093/nar/gkr1013 doi: 10.1093/nar/gkr1013
|
| [25] |
M. Johnson, I. Zaretskaya, Y. Raytselis, Y. Merezhuk, S. McGinnis, T. L. Madden, NCBI BLAST: a better web interface, Nucleic Acids Res., 36 (2008), W5–W9. https://doi.org/10.1093/nar/gkn201 doi: 10.1093/nar/gkn201
|
| [26] |
C. von Mering, M. Huynen, D. Jaeggi, S. Schmidt, P. Bork, B. Snel, STRING: a database of predicted functional associations between proteins, Nucleic Acids Res., 31 (2003), 258–261. https://doi.org/10.1093/nar/gkg034 doi: 10.1093/nar/gkg034
|
| [27] |
D. W. Huang, B. T. Sherman, R. A. Lempicki, Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources, Nat. Protoc., 4 (2009), 44–57. https://doi.org/10.1038/nprot.2008.211 doi: 10.1038/nprot.2008.211
|
| [28] |
T. Shibata, H. Aburatani, Exploration of liver cancer genomes, Nat. Rev. Gastro. Hepat., 11 (2014), 340–349. https://doi.org/10.1038/nrgastro.2014.6 doi: 10.1038/nrgastro.2014.6
|
| [29] |
L. Wang, K. Yan, X. He, H. Zhu, J. Song, S. Chen, et al., LRP1B or TP53 mutations are associated with higher tumor mutational burden and worse survival in hepatocellular carcinoma, J. Cancer, 12 (2021), 217–223. https://doi.org/10.7150/jca.48983 doi: 10.7150/jca.48983
|
| [30] |
B. Wu, R. Liu, PAQR4 promotes cell proliferation and metastasis through the CDK4-pRB-E2F1 pathway in non-small-cell lung cancer, Onco Targets Ther., 12 (2019), 3625–3633. https://doi.org/10.2147/OTT.S181432 doi: 10.2147/OTT.S181432
|
| [31] |
R. P. Kuiper, M. J. L. Ligtenberg, N. Hoogerbrugge, A. G. Kessel, Germline copy number variation and cancer risk, Curr. Opin. Genet. Dev., 20 (2010), 282–289. https://doi.org/10.1016/j.gde.2010.03.005 doi: 10.1016/j.gde.2010.03.005
|
| [32] |
A. Eden, F. Gaudet, A. Waghmare, R. Jaenisch, Chromosomal instability and tumors promoted by DNA hypomethylation, Science, 300 (2003), 455. https://doi.org/10.1126/science.1083557 doi: 10.1126/science.1083557
|
| [33] |
T. Yang, X. Liu, S.K. Kumar, F. Jin, Y. Dai, Decoding DNA methylation in epigenetics of multiple myeloma, Blood Rev., 51 (2021), 100872. https://doi.org/10.1016/j.blre.2021.100872 doi: 10.1016/j.blre.2021.100872
|
| [34] |
N. Lu, J. Liu, C. Ji, Y. Wang, Z. Wu, S. Yuan, et al., MiRNA based tumor mutation burden diagnostic and prognostic prediction models for endometrial cancer, Bioengineered, 12 (2021), 3603–3620. https://doi.org/10.1080/21655979.2021.1947940 doi: 10.1080/21655979.2021.1947940
|
| [35] |
M. I. Almeida, R. M. Reis, G. A. Calin, MicroRNA history: discovery, recent applications, and next frontiers, Mutat. Res., 717 (2011), 1–8. https://doi.org/10.1016/j.mrfmmm.2011.03.009 doi: 10.1016/j.mrfmmm.2011.03.009
|
| [36] |
C. Tang, Y. Wu, X. Wang, K. Chen, Z. Tang, X. Guo, LncRNA MAFG-AS1 regulates miR-125b-5p/SphK1 axis to promote the proliferation, migration, and invasion of bladder cancer cells, Hum. Cell, 34 (2021), 588–597. https://doi.org/10.1007/s13577-020-00470-3 doi: 10.1007/s13577-020-00470-3
|
| [37] |
B. Hu, X. B. Yang, X. Yang, X. T. Sang, LncRNA CYTOR affects the proliferation, cell cycle and apoptosis of hepatocellular carcinoma cells by regulating the miR-125b-5p/KIAA1522 axis, Aging (Albany NY), 13 (2020), 2626–2639. https://doi.org/10.18632/aging.202306 doi: 10.18632/aging.202306
|
| [38] |
Y. Li, Y. Wang, H. Fan, Z. Zhang, N. Li, miR-125b-5p inhibits breast cancer cell proliferation, migration and invasion by targeting KIAA1522, Biochem. Bioph. Res. Commun., 504 (2018), 277–282. https://doi.org/10.1016/j.bbrc.2018.08.172 doi: 10.1016/j.bbrc.2018.08.172
|
| [39] |
J. Nyqvist, A. Kovács, Z. Einbeigi, P. Karlsson, E. Forssell-Aronsson, K. Helou, et al., Genetic alterations associated with multiple primary malignancies, Cancer Med., 10 (2021), 4465–4477. https://doi.org/10.1002/cam4.3975 doi: 10.1002/cam4.3975
|
| [40] |
K. Shimada, K. Yoshida, Y. Suzuki, C. Iriyama, Y. Inoue, M. Sanada, et al., Frequent genetic alterations in immune checkpoint-related genes in intravascular large B-cell lymphoma, Blood, 137 (2021), 1491–1502. https://doi.org/10.1182/blood.2020007245 doi: 10.1182/blood.2020007245
|
| [41] |
L. Wang, R. Zhang, X. You, H. Zhang, S. Wei, T. Cheng, et al., The steady-state level of CDK4 protein is regulated by antagonistic actions between PAQR4 and SKP2 and involved in tumorigenesis, J. Mol. Cell Biol, . 9 (2017), 409–421. https://doi.org/10.1093/jmcb/mjx028 doi: 10.1093/jmcb/mjx028
|
| [42] |
H. Zhang, R. Han, Z. Q. Ling, F. Zhang, Y. Hou, X. You, et al., PAQR4 has a tumorigenic effect in human breast cancers in association with reduced CDK4 degradation, Carcinogenesis, 39 (2018), 439–446. https://doi.org/10.1093/carcin/bgx143 doi: 10.1093/carcin/bgx143
|
| [43] |
M. Guo, N. Li, J. Zheng, W. Wang, Y. Wu, X. Han, et al., Epigenetic regulation of hepatocellular carcinoma progression through the mTOR signaling pathway, Can. J. Gastroenterol. Hepatol., 2021 (2021), 5596712. https://doi.org/10.1155/2021/5596712 doi: 10.1155/2021/5596712
|
| [44] |
H. Moon, S. W. Ro, MAPK/ERK signaling pathway in hepatocellular carcinoma, Cancers (Basel), 13 (2021), 3026. https://doi.org/10.3390/cancers13123026 doi: 10.3390/cancers13123026
|
| [45] |
L. J. Liu, S. X. Xie, Y. T. Chen, J. L. Xue, C. J. Zhang, F. Zhu, Aberrant regulation of Wnt signaling in hepatocellular carcinoma, World J. Gastroenterol., 22 (2016), 7486–7499. https://doi.org/10.3748/wjg.v22.i33.7486 doi: 10.3748/wjg.v22.i33.7486
|
| [46] |
Y. Feng, T. Sun, Y. Yu, Y. Gao, X. Wang, Z. Chen, MicroRNA-370 inhibits the proliferation, invasion and EMT of gastric cancer cells by directly targeting PAQR4, J. Pharmacol. Sci, . 138 (2018), 96–106. https://doi.org/10.1016/j.jphs.2018.08.004 doi: 10.1016/j.jphs.2018.08.004
|
| [47] |
A. Del Moral-Morales, J. C. González-Orozco, J. M. Capetillo-Velázquez, A. G. Piña-Medina, I. Camacho-Arroyo, The role of mPRδ and mPRε in human glioblastoma cells: expression, hormonal regulation, and possible clinical outcome, Horm Cancer, 11 (2020), 117–127. https://doi.org/10.1007/s12672-020-00381-7 doi: 10.1007/s12672-020-00381-7
|















Figures(16) / Tables(2)

Caihao Qu, Tengda Ma, Xin YAN, Xiaomei Li, Yumin Li. Overexpressed PAQR4 predicts poor overall survival and construction of a prognostic nomogram based on PAQR family for hepatocellular carcinoma[J]. Mathematical Biosciences and Engineering, 2022, 19(3): 3069-3090. doi: 10.3934/mbe.2022142







 DownLoad:
DownLoad: