Citation: Dongmei Wu, Hao Wang, Sanling Yuan. Stochastic sensitivity analysis of noise-induced transitions in a predator-prey model with environmental toxins[J]. Mathematical Biosciences and Engineering, 2019, 16(4): 2141-2153. doi: 10.3934/mbe.2019104

| [1] | Q. Huang, H. Wang and M. A. Lewis, The impact of environmental toxins on predator-prey dynamics, J. Theor. Biol., 378 (2015), 12–30. |
| [2] | R. A. Pastorok, S. M. Bartell and S. Ferson, et al., Ecological modeling in risk assessment: chemical effects on populations, ecosystems, and landscapes, Lewis Publishers, Boca Raton, FL, USA, 2001. |
| [3] | S. M. Bartell, R. A. Pastorok and H. R. Akcakaya, et al., Realism and relevance of ecological models used in chemical risk assessment, Hum. Ecol. Risk Assess., 9 (2003), 907–938. |
| [4] | N. Galic, U. Hommen and J. H. Baveco, et al., Potential application of population models in the European ecological risk assessment of chemicals. II: review of models and their potential to address environmental protection aims, Integr. Environ. Assess. Manag., 6 (2010), 338–360. |
| [5] | Y. Zhao, S. Yuan and T. Zhang, The stationary distribution and ergodicity of a stochastic phytoplankton allelopathy model under regime switching, Commun. Nonlinear Sci. Numer. Simtlat., 37 (2016), 131–142. |
| [6] | X. Yu, S. Yuan and T. Zhang, The effects of toxin-producing phytoplankton and environmental fluctuations on the planktonic blooms, Nonlinear Dyn., 91 (2018), 1653–1668. |
| [7] | H. R. Thieme, Mathematics in population biology, Princeton Series in Theoretical and Computational Biology, 2003. |
| [8] | Q. Huang, L. Parshotham and H. Wang, et al., A model for the impact of contaminants on fish population dynamics, J. Theor. Biol., 334 (2013), 71–79. |
| [9] | B. C. Kelly, M. G. Ikonomou and J. D. Blair, et al., Food webspecific biomagnification of persistent organic pollutants, Science, 317 (2007), 236–239. |
| [10] | R. V. Thomann, Bioaccumulation model of organic chemical distribution in aquatic food chains, Environ. Sci. Technol., 23 (1989), 699–707. |
| [11] | V. S. Anishchenko, V. Astakhov and A. Neiman, et al., Nonlinear dynamics of chaotic and stochastic systems, Springer, Berlin, 2007. |
| [12] | A. N. Pisarchik and U. Feudel, Control of multistability, Phys. Rep., 540 (2014), 167–218. |
| [13] | C. Xu, S. Yuan and T. Zhang, Stochastic sensitivity analysis for a competitive turbidostat model with inhibitory nutrients, Int. J. Bifurcat. Chaos, 8 (2016), 1440020. |
| [14] | C. Xu, S. Yuan and T. Zhang, Sensitivity analysis and feedback control of noise-induced extinction for competition chemostat model with mutualism, Physica A, 505 (2018), 891–902. |
| [15] | C. Kurrer and K. Schulten, Effect of noise and perturbations on limit cycle systems, Physica D, 50 (1991), 311–320. |
| [16] | F. Baras, Lecture notes in physics springer-verlag, New York, 484 (1997), 167–178. |
| [17] | L. Gammaitoni, P. H. anggi and P. Jung, et al., Stochastic resonance, Rev. Mod. Phys., 70 (1999), 223. |
| [18] | M. D. McDonnell, N. G. Stocks and C. E. M. Pearce, et al., Stochastic resonance: from suprathreshold stochastic resonance to stochastic signal quantization cambridge, University Press, New York, 2008. |
| [19] | W. Horsthemke and R. Lefever, Noise-induced transitions, Springer, Berlin, 1984. |
| [20] | K. Matsumoto and I. Tsuda, Noise-induced order, J. Stat. Phys., 33 (1983), 757. |
| [21] | F. Gassmann, Noise-induced chaos-order transitions, Phys. Rev. E, 55 (1997), 2215–2221. |
| [22] | J. Gao, S. Hwang and J. Liu, When can noise induce chaos? Phys. Rev. Lett., 82 (1999), 1132– 1135. |
| [23] | M. A. Zaks, X. Sailer and L. Schimansky-Geier, et al., Noise induced complexity: From subthreshold oscillations to spiking in coupled excitable systems, Chaos, 15 (2005), 026117. |
| [24] | C. Xu and S. Yuan, An analogue of break-even concentration in a simple stochastic chemostat model, Appl. Math. Lett., 48 (2015), 62–68. |
| [25] | Y. Zhao, S. Yuan and J. Ma, Survival and stationary distribution analysis of a stochastic competitive model of three species in a polluted environment, B. Math. Biol., 77 (2015), 1285–1326. |
| [26] | Y. Zhao, S. Yuan and T. Zhang, Stochastic periodic solution of a non-autonomous toxic-producing phytoplankton allelopathy model with environmental fluctuation, Commun. Nonlinear Sci. Numer. Simtlat., 44 (2017), 266–276. |
| [27] | X. Yu, S. Yuan and T. Zhang, Persistence and ergodicity of a stochastic single species model with Allee effect under regime switching, Commun. Nonlinear Sci. Numer. Simtlat., 59 (2018), 359–374. |
| [28] | X. Yu, S. Yuan and T. Zhang, About the optimal harvesting of a fuzzy predator-prey system: A bioeconomic model incorporating a prey refuge and predator mutual interference, Nonlinear Dyn., DOI: 10.1007/s11071-018-4480-y. |
| [29] | C. Xu and S. Yuan, Competition in the chemostat: a stochastic multi-species model and its asymptotic behavior, Math. Biosci., 280 (2016), 1–9. |
| [30] | S. Kim, S. H. Park and C. S. Ryu, Colored-noise-induced multistability in nonequilibrium phase transitions, Phys. Rev. E, 58 (1998), 7994–7997. |
| [31] | S. Kraut and U. Feudel, Multistability, noise, and attractor hopping: the crucial role of chaotic saddles, Phys. Rev. E, 66 (2002), 015207(R). |
| [32] | S. L. T. de Souza, A. M. Batista and I. L. Caldas, et al., Noise-induced basin hopping in a vibro-impact system, Chaos Solit. Fract., 32 (2007), 758–767. |
| [33] | M. I. Dykman, R. Mannella and P. V. E. McClintock, et al., Fluctuation-induced transitions between periodic attractors: Observation of supernarrow spectral peaks near a kinetic phase transition, Phys. Rev. Lett., 66 (1990), 48. |
| [34] | M. I. Dykman, D. G. Luchinsky and R. Mannella, et al., Simulation of critical phenomena in nonlinear optical systems, Phys. Rev. E, 49 (1994), 1198. |
| [35] | I. Bashkirtseva, L. Ryashko and I. Tsvetkov, Sensitivity analysis of stochastic equilibria and cycles for the discrete dynamic systems, Dyn. Contin. Discrete Impuls. Syst. Ser. A Math. Anal., 17 (2010), 501–515. |
| [36] | I. Bashkirtseva, T. Ryazanova and L. Ryashko, Confidence domains in the analysis of noiseinduced transition to chaos for Goodwin model of business cycles, Int. J. Bifurc. Chaos., 24 (2014), 1440020. |
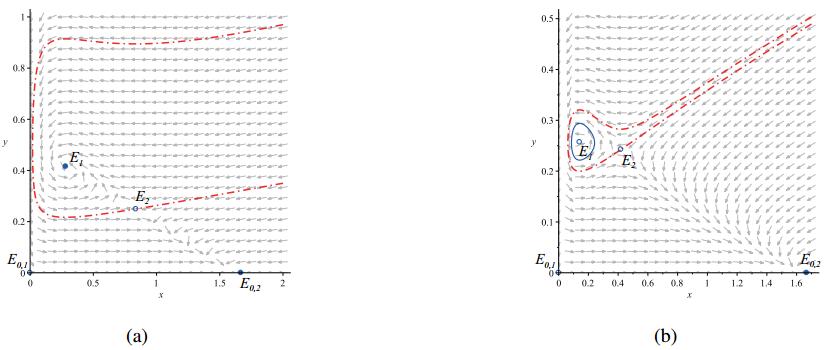
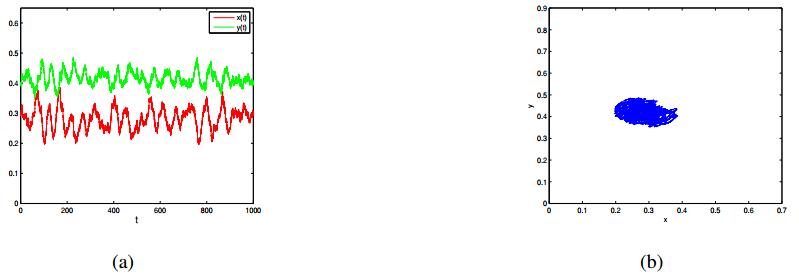




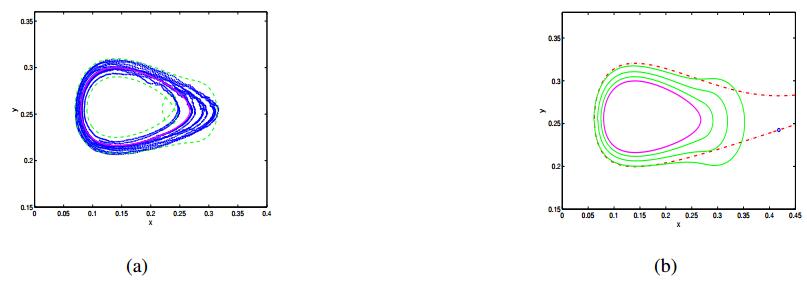
Figures(7)

Dongmei Wu, Hao Wang, Sanling Yuan. Stochastic sensitivity analysis of noise-induced transitions in a predator-prey model with environmental toxins[J]. Mathematical Biosciences and Engineering, 2019, 16(4): 2141-2153. doi: 10.3934/mbe.2019104







 DownLoad:
DownLoad: