In this paper, we investigate the dynamical properties of a stochastic predator-prey model with a fear effect. We also introduce infectious disease factors into prey populations and distinguish prey populations into susceptible prey and infected prey populations. Then, we discuss the effect of Lévy noise on the population considering extreme environmental situations. First of all, we prove the existence of a unique global positive solution for this system. Second, we demonstrate the conditions for the extinction of three populations. Under the conditions that infectious diseases are effectively prevented, the conditions for the existence and extinction of susceptible prey populations and predator populations are explored. Third, the stochastic ultimate boundedness of system and the ergodic stationary distribution without Lévy noise are also demonstrated. Finally, we use numerical simulations to verify the conclusions obtained and summarize the work of the paper.
Citation: Xueqing He, Ming Liu, Xiaofeng Xu. Analysis of stochastic disease including predator-prey model with fear factor and Lévy jump[J]. Mathematical Biosciences and Engineering, 2023, 20(2): 1750-1773. doi: 10.3934/mbe.2023080
In this paper, we investigate the dynamical properties of a stochastic predator-prey model with a fear effect. We also introduce infectious disease factors into prey populations and distinguish prey populations into susceptible prey and infected prey populations. Then, we discuss the effect of Lévy noise on the population considering extreme environmental situations. First of all, we prove the existence of a unique global positive solution for this system. Second, we demonstrate the conditions for the extinction of three populations. Under the conditions that infectious diseases are effectively prevented, the conditions for the existence and extinction of susceptible prey populations and predator populations are explored. Third, the stochastic ultimate boundedness of system and the ergodic stationary distribution without Lévy noise are also demonstrated. Finally, we use numerical simulations to verify the conclusions obtained and summarize the work of the paper.

| [1] |
P. Naik, Z. Eskandari, Z. Avazzadeh, J. Zu, Multiple bifurcations of a discrete-time prey-predator model with mixed functional response, Int. J. Bifurcat. Chaos, 32 (2022). https://doi.org/10.1142/S021812742250050X doi: 10.1142/S021812742250050X

|
| [2] |
J. Beddington, Mutual interference between parasites or predators and its effect on searching efficiency, J. Animal. Ecol., 44 (1975), 331–340. https://doi.org/10.2307/3866 doi: 10.2307/3866
|
| [3] |
D. L. DeAngelis, R. A. Goldstein, R. V. ONeill, A model for trophic interaction, Ecology, 56 (1975), 881–892. https://doi.org/10.2307/1936298 doi: 10.2307/1936298
|
| [4] |
P. Naik, Z. Eskandari, M. Yavuz, J. Zu, Complex dynamics of a discrete-time Bazykin-Berezovskaya prey-predator model with a strong Allee effect, J. Comput. Appl. Math., 413 (2022). https://doi.org/10.1016/j.cam.2022.114401 doi: 10.1016/j.cam.2022.114401
|
| [5] | A. Lotka, Elements of Physical Biology, Williams and Wilkins, Galtimore, 1925. https://doi.org/10.1038/116461b0 |
| [6] |
V. Voltera, Variazioni e fluttuazioni del numero di individui in specie animali conviventi, Mem. Accd. Lincei., 6 (1926), 31–113. https://doi.org/10.1038/118558a0 doi: 10.1038/118558a0
|
| [7] |
P. Leslie, Some further notes on the use of matrices in population mathematic, Biometrica, 35 (1948), 213–245. https://doi.org/10.1093/biomet/35.3-4.213 doi: 10.1093/biomet/35.3-4.213
|
| [8] |
P. Leslie, J. Gower, The properties of a stochastic model for the predator-prey type of interaction between two species, Biometrica, 47 (1960), 219–234. https://doi.org/10.1093/biomet/47.3-4.219 doi: 10.1093/biomet/47.3-4.219
|
| [9] |
M. Aziz-Alaoui, M. Okiye, Boundedness and global stability for a predator-prey model with modified Leslie-Gower and Holling-type Ⅱ schemes, Appl. Math. Lett., 16 (2003), 1069–1075. https://doi.org/10.1016/S0893-9659(03)90096-6 doi: 10.1016/S0893-9659(03)90096-6
|
| [10] |
A. Kumar, Nilam, Stability of a time delayed SIR epidemic model along with nonlinear incidence rate and Holling type-Ⅱ treatment rate, Int. J. Comput. Meth., 15 (2018). https://doi.org/10.1142/S021987621850055X doi: 10.1142/S021987621850055X
|
| [11] |
S. Rezapour, S. Etemad, H. Mohammadi, A mathematical analysis of a system of Caputo-Fabrizio fractional differential equations for the anthrax disease model in animals, Adv. Differ. Equations, 2020 (2020), 418. https://doi.org/10.1186/s13662-020-02937-x doi: 10.1186/s13662-020-02937-x
|
| [12] |
H. Mohammadi, S. Kumar, S. Rezapour, S. Etemad, A theoretical study of the Caputo-Fabrizio fractional modeling for hearing loss due to Mumps virus with optimal control, Chaos, Solitons Fractals, 144 (2021). https://doi.org/10.1016/j.chaos.2021.110668 doi: 10.1016/j.chaos.2021.110668
|
| [13] |
N. Tuan, H. Mohammadi, S. Rezapour, A mathematical model for COVID-19 transmission by using the Caputo fractional derivative, Chaos, Solitons Fractals, 140 (2021). https://doi.org/10.1016/j.chaos.2020.110107 doi: 10.1016/j.chaos.2020.110107
|
| [14] |
D. Baleanu, H. Mohammadi, S. Rezapour, Analysis of the model of HIV-1 infection of CD4(+) T-cell with a new approach of fractional derivative, Adv. Differ. Equations, 2020 (2020), 71. https://doi.org/10.1186/s13662-020-02544-w doi: 10.1186/s13662-020-02544-w
|
| [15] |
Y. Liu, M. liu, X. Xu, Dynamics analysis of stochastic modified Leslie-Gower model with time-delay and Michaelis-Menten type prey harvest, J. Appl. Math. Comput., 68 (2020), 2097–2124. https://doi.org/10.1007/s12190-021-01612-y doi: 10.1007/s12190-021-01612-y
|
| [16] |
D. Zhou, M. Liu, Z. Liu, Persistence and extinction of a stochastic predator-prey model with modified Leslie-Gower and Holling-type Ⅱ schemes, Adv. Differ. Equations, 179 (2020). https://doi.org/10.1186/s13662-020-02642-9 doi: 10.1186/s13662-020-02642-9
|
| [17] |
A. Majumder, D. Adak, N. Bairagi, Persistence and extinction of species in a disease-induced ecological system under environmental stochasticity, Phys. Rev. E., 103 (2021). https://doi.org/10.1103/PhysRevE.103.032412 doi: 10.1103/PhysRevE.103.032412
|
| [18] |
L. Zanette, A. White, M. Allen, M. Clinchy, Perceived predation risk reduces the number of offspring songbirds produce per year, Science, 334 (2011), 1398–1401. https://doi.org/10.1126/science.1210908 doi: 10.1126/science.1210908
|
| [19] |
J. Roy, D. Barman, S. Alam, Role of fear in a predator-prey system with ratio-dependent functional response in deterministic and stochastic environment, Biosystems, 197 (2020). https://doi.org/10.1016/j.biosystems.2020.104176 doi: 10.1016/j.biosystems.2020.104176
|
| [20] |
H. Zhang, Y. Cai, S. Fu, W. Wang, Impact of the fear effect in a prey-predator model incorporating a prey refuge, Appl. Math. Comput., 356 (2019), 328–337. https://doi.org/10.1016/j.amc.2019.03.034 doi: 10.1016/j.amc.2019.03.034
|
| [21] |
K. Sarkara, S. Khajanchi, Impact of fear effect on the growth of prey in a predator-prey interaction model, Ecol. Complex., 42 (2020). https://doi.org/10.1016/j.ecocom.2020.100826 doi: 10.1016/j.ecocom.2020.100826
|
| [22] |
H. Qi, X. Meng, Threshold behavior of a stochastic predator-prey system with prey refuge and fear effect, Appl. Math. Lett., 113 (2021). https://doi.org/10.1016/j.aml.2020.106846 doi: 10.1016/j.aml.2020.106846
|
| [23] |
J. Bao, X. Mao, G. Yin, C. Yuan, Competitive Lotka-Volterra population dynamics with jumps, Nonlinear Anal: Theory, Methods Appl., 74 (2011), 6601–6616. https://doi.org/10.1016/j.na.2011.06.043 doi: 10.1016/j.na.2011.06.043
|
| [24] |
J. Bao, C. Yuan, Stochastic population dynamics driven by Lévy noise, J. Math. Anal. Appl., 391 (2012), 363–375. https://doi.org/10.1016/j.jmaa.2012.02.043 doi: 10.1016/j.jmaa.2012.02.043
|
| [25] |
R. Wu, K. Wang, Population dynamical behaviors of stochastic logistic system with jumps, Turk. J. Math., 38 (2014), 935–948. https://doi.org/10.3906/mat-1307-25 doi: 10.3906/mat-1307-25
|
| [26] | R. Wu, X. Zou, K. Wang, Dynamics of Logistic system driven Lévy noise under regime switching, Electron. J. Differ. Equations, 76 (2014), 1–16. |
| [27] |
S. Zhang, X. Meng, T. Feng, T. Zhang, Dynamics analysis and numerical simulations of a stochastic non-autonomous predator-prey system with impulsive effects, Nonlinear Anal.: Hybrid Syst., 26 (2017), 19–37. https://doi.org/10.1016/j.nahs.2017.04.003 doi: 10.1016/j.nahs.2017.04.003
|
| [28] |
R. Lipster, A strong law of large numbers for local martingales, Stochastics, 3 (1980), 217–228. https://doi.org/110.1080/17442508008833146 doi: 10.1080/17442508008833146
|
| [29] |
R. Khasminskii, F. Klebaner, Long term behavior of solutions of the Lotka-Volterra system under small random perturbations, Ann. Appl. Probab., 11 (2001), 952–963. https://doi.org/10.1214/aoap/1015345354 doi: 10.1214/aoap/1015345354
|
| [30] |
M. Liu, K. Wang, Survival analysis of a stochastic cooperation system in a polluted environment, J. Biol. Syst., 19 (2011), 183–204. https://doi.org/10.1142/S0218339011003877 doi: 10.1142/S0218339011003877
|
| [31] |
X. Li, A. Alison, D. Jiang, X. Mao, Sufficient and necessary conditions of stochastic permanence and extinction for stochastic logistic populations under regimes witching, J. Math. Anal. Appl., 376 (2011), 11–28. https://doi.org/10.1016/j.jmaa.2010.10.053 doi: 10.1016/j.jmaa.2010.10.053
|
| [32] | R. Khasminskii, Stochastic Stability of Differential Equations, in Stochastic Modeling and Applied Probability, Springer Berlin, Heidelberg, (2012), 99–136. https://doi.org/10.1007/978-3-642-23280-0 |
| [33] |
Q. Liu, D. Jiang, N. Shi, T. Hayat, A. Alsaedi, Dynamical behavior of a stochastic HBV infection model with logistic hepatocyte growth, Acta. Math. Sci., 37 (2017), 927–940. https://doi.org/10.1016/S0252-9602(17)30048-6 doi: 10.1016/S0252-9602(17)30048-6
|
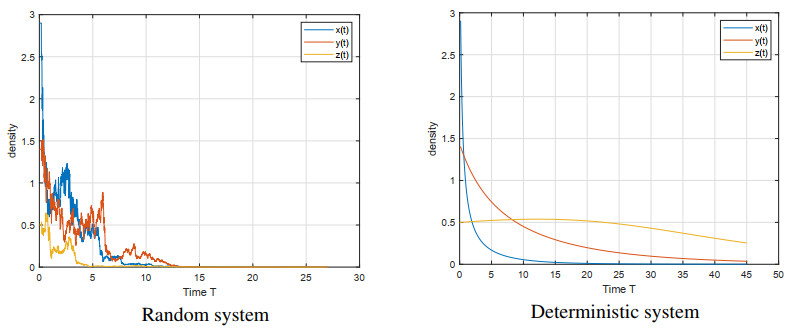
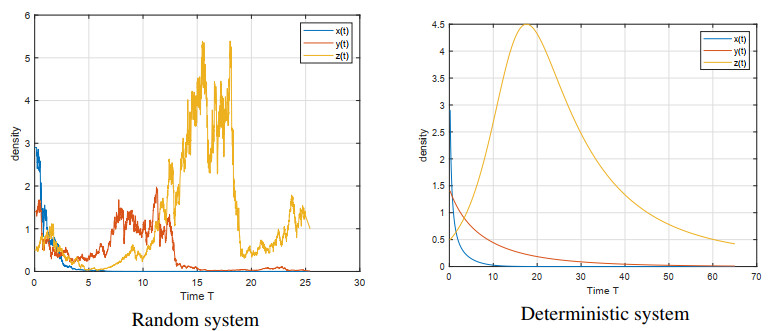
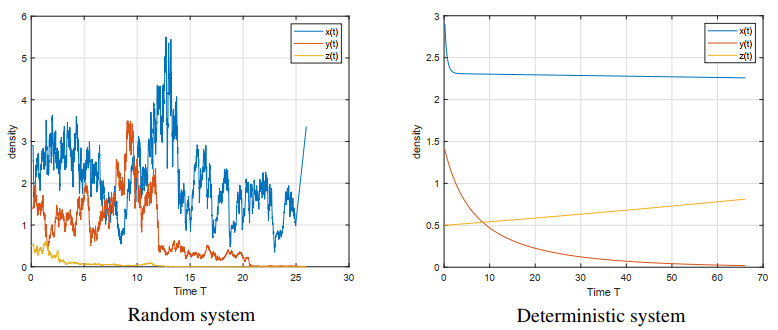
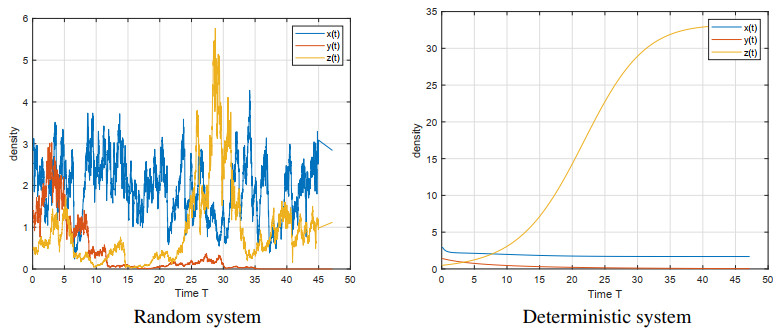
Figures(5)

Xueqing He, Ming Liu, Xiaofeng Xu. Analysis of stochastic disease including predator-prey model with fear factor and Lévy jump[J]. Mathematical Biosciences and Engineering, 2023, 20(2): 1750-1773. doi: 10.3934/mbe.2023080







 DownLoad:
DownLoad: