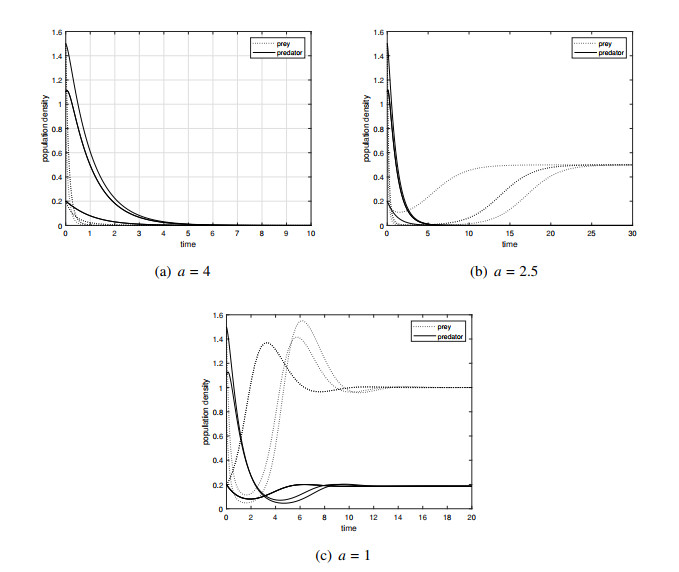
We have combined cooperative hunting, inspired by recent experimental studies on birds and vertebrates, to develop a predator-prey model in which the fear effect simultaneously influences the birth and mortality rates of the prey. This differs significantly from the fear effect described by most scholars. We have made a comprehensive analysis of the dynamics of the model and obtained some new conclusions. The results indicate that both fear and cooperative hunting can be a stable or unstable force in the system. The fear can increase the density of the prey, which is different from the results of all previous scholars, and is a new discovery in our study of the fear effect. Another new finding is that fear has an opposite effect on the densities of two species, which is different from the results of most other scholars in that fear synchronously reduces the densities of both species. Numerical simulations have also revealed that the fear effect extends the time required for the population to reach its survival state and accelerates the process of population extinction.
Citation: Yalong Xue, Fengde Chen, Xiangdong Xie, Shengjiang Chen. An analysis of a predator-prey model in which fear reduces prey birth and death rates[J]. AIMS Mathematics, 2024, 9(5): 12906-12927. doi: 10.3934/math.2024630
We have combined cooperative hunting, inspired by recent experimental studies on birds and vertebrates, to develop a predator-prey model in which the fear effect simultaneously influences the birth and mortality rates of the prey. This differs significantly from the fear effect described by most scholars. We have made a comprehensive analysis of the dynamics of the model and obtained some new conclusions. The results indicate that both fear and cooperative hunting can be a stable or unstable force in the system. The fear can increase the density of the prey, which is different from the results of all previous scholars, and is a new discovery in our study of the fear effect. Another new finding is that fear has an opposite effect on the densities of two species, which is different from the results of most other scholars in that fear synchronously reduces the densities of both species. Numerical simulations have also revealed that the fear effect extends the time required for the population to reach its survival state and accelerates the process of population extinction.

| [1] |
R. H. Hering, Oscillations in Lotka-Volterra systems of chemical reactions, J. Math. Chem., 5 (1990), 197–202. https://doi.org/10.1007/BF01166429 doi: 10.1007/BF01166429

|
| [2] | F. H. Busse, Transition to turbulence via the statistical limit cycle route, In: Chaos and order in nature, Berlin, Heidelberg: Springer, 1981, 36–44. https://doi.org/10.1007/978-3-642-68304-6_4 |
| [3] |
S. Solomon, P. Richmond, Stable power laws in variable economies; Lotka-Volterra implies Pareto-Zipf, Eur. Phys. J. B., 27 (2002), 257–261. https://doi.org/10.1140/epjb/e20020152 doi: 10.1140/epjb/e20020152
|
| [4] |
G. Laval, R. Pellat, M. Perulli, Study of the disintegration of Langmuir waves, Plasma Phys., 11 (1969), 579–588. https://doi.org/10.1088/0032-1028/11/7/003 doi: 10.1088/0032-1028/11/7/003
|
| [5] |
Z. Wang, M. Jusup, L. Shi, J. H. Lee, Y. Iwasa, S. Boccaletti, Exploiting a cognitive bias promotes cooperation in social dilemma experiments, Nat. Commun., 9 (2018), 2954. https://doi.org/10.1038/s41467-018-05259-5 doi: 10.1038/s41467-018-05259-5
|
| [6] |
Z. Wang, M. Jusup, H. Guo, L. Shi, S. Geček, M. Anand, et al., Communicating sentiment and outlook reverses inaction against collective risks, PNAS, 117 (2020), 17650–17655. https://doi.org/10.1073/pnas.1922345117 doi: 10.1073/pnas.1922345117
|
| [7] |
N. N. Pelen, On the dynamics of impulsive predator-prey systems with Beddington-Deangelis-type functional response, Ukr. Math. J., 73 (2021), 610–634. https://doi.org/10.1007/s11253-021-01947-6 doi: 10.1007/s11253-021-01947-6
|
| [8] |
T. T. Ma, X. Z. Men, T. Hayat, A. Hobiny, Hopf bifurcation induced by time delay and influence of Allee effect in a diffusive predator-prey system with herd behavior and prey chemotaxis, Nonlinear Dyn., 108 (2022), 4581–4598. https://doi.org/10.1007/s11071-022-07401-x doi: 10.1007/s11071-022-07401-x
|
| [9] |
A. Mezouaghi, S. Djilali, S. Bentout, K. Biroud, Bifurcation analysis of a diffusive predator-prey model with prey social behavior and predator harvesting, Math. Methods Appl. Sci., 45 (2022), 718–731. https://doi.org/10.1002/mma.7807 doi: 10.1002/mma.7807
|
| [10] |
M. S. Islam, N. Sk, S. Sarwardi, Deterministic and stochastic study of an eco-epidemic predator-prey model with nonlinear prey refuge and predator harvesting, Eur. Phys. J. Plus, 138 (2023), 851. https://doi.org/10.1140/epjp/s13360-023-04476-2 doi: 10.1140/epjp/s13360-023-04476-2
|
| [11] |
Y. Y. Dong, G. Gao, S. B. Li, Coexistence states for a prey-predator model with cross-diffusion, J. Math. Anal. Appl., 535 (2024), 128106. https://doi.org/10.1016/j.jmaa.2024.128106 doi: 10.1016/j.jmaa.2024.128106
|
| [12] |
R. Cherniha, V. Davydovych, Symmetries and exact solutions of the diffusive Holling-Tanner prey-predator model, Acta Appl. Math., 187 (2023), 8. https://doi.org/10.1007/s10440-023-00600-7 doi: 10.1007/s10440-023-00600-7
|
| [13] |
L. Y. Liu, C. Y. Yang, A free boundary problem for a ratio-dependent predator-prey system, Z. Angew. Math. Phys., 74 (2023), 69. https://doi.org/10.1007/s00033-023-01957-3 doi: 10.1007/s00033-023-01957-3
|
| [14] |
C. Liu, L. L. Chang, Y. Huang, Z. Wang, Turing patterns in a predator-prey model on complex networks, Nonlinear Dyn., 99 (2020), 3313–3322. https://doi.org/10.1007/s11071-019-05460-1 doi: 10.1007/s11071-019-05460-1
|
| [15] |
A. Sha, D. S. Mandal, A. Chekroun, Impact of prey refuge in a diffusive prey predator model with prey harvesting, mutually interfering predator and additional food for predator, Int. J. Appl. Comput. Math., 9 (2023), 56. https://doi.org/10.1007/s40819-023-01546-y doi: 10.1007/s40819-023-01546-y
|
| [16] |
S. Djilali, S. Bentout, Pattern formations of a delayed diffusive predator-prey model with predator harvesting and prey social behavior, Math. Methods Appl. Sci., 44 (2021), 9128–9142. https://doi.org/10.1002/mma.7340 doi: 10.1002/mma.7340
|
| [17] |
L. Y. Zanette, A. F. White, M. C. Allen, M. Clinchy, Perceived predation risk reduces the number of offspring songbirds produce per year, Science, 334 (2011), 1398–1401. https://doi.org/10.1126/science.1210908 doi: 10.1126/science.1210908
|
| [18] |
S. Creel, D. Christianson, S. Liley, J. A. Winnie Jr., Predation risk affects reproductive physiology and demography of elk, Science, 315 (2007), 960. https://doi.org/10.1126/science.1135918 doi: 10.1126/science.1135918
|
| [19] |
M. J. Sheriff, C. J. Krebs, R. Boonstra, The sensitive hare: sublethal effects of predator stress on reproduction in snowshoe hares, J. Anim. Ecol., 78 (2009), 1249–1258. https://doi.org/10.1111/j.1365-2656.2009.01552.x doi: 10.1111/j.1365-2656.2009.01552.x
|
| [20] |
A. J. Wirsing, W. J. Ripple, A comparison of shark and wolf research reveals similar behavioral responses by prey, Front. Ecol. Environ., 9 (2011), 335–341. https://doi.org/10.1890/090226 doi: 10.1890/090226
|
| [21] |
X. Y. Wang, L. Zanette, X. F. Zou, Modelling the fear effect in predator-prey interactions, J. Math. Biol., 73 (2016), 1179–1204. https://doi.org/10.1007/s00285-016-0989-1 doi: 10.1007/s00285-016-0989-1
|
| [22] |
Z. L. Zhu, R. X. Wu, L. Y. Lai, X. Q. Yu, The influence of fear effect to the Lotka-Volterra predator-prey system with predator has other food resource, Adv. Differ. Equ., 2020 (2020), 237. https://doi.org/10.1186/s13662-020-02612-1 doi: 10.1186/s13662-020-02612-1
|
| [23] |
S. Pal, N. Pal, S. Samanta, J. Chattopadhyay, Fear effect in prey and hunting cooperation among predators in a Leslie-Gower model, Math. Biosci. Eng., 16 (2019), 5146–5179. https://doi.org/10.3934/mbe.2019258 doi: 10.3934/mbe.2019258
|
| [24] |
S. K. Sasmal, Population dynamics with multiple Allee effect induced by fear factors–A mathematical study on prey-predator interactions, Appl. Math. Model., 64 (2018), 1–14. https://doi.org/10.1016/j.apm.2018.07.021 doi: 10.1016/j.apm.2018.07.021
|
| [25] |
B. Dennis, Allee effects: population growth, critical density and the chance of extinction, Nat. Resour. Model., 3 (1989), 481–538. https://doi.org/10.1111/j.1939-7445.1989.tb00119.x doi: 10.1111/j.1939-7445.1989.tb00119.x
|
| [26] |
L. Y. Lai, Z. L. Zhu, F. D. Chen, Stability and bifurcation in a predator-prey model with the additive Allee effect and the fear effect, Mathematics, 8 (2020), 1–21. https://doi.org/10.3390/math8081280 doi: 10.3390/math8081280
|
| [27] |
H. S. Zhang, Y. L. Cai, S. M. Fu, W. M. Wang, Impact of the fear effect in a prey-predator model incorporating a prey refuge, Appl. Math. Comput., 356 (2019), 328–337. https://doi.org/10.1016/j.amc.2019.03.034 doi: 10.1016/j.amc.2019.03.034
|
| [28] |
M. F. Carfora, I. Torcicollo, Cross-diffusion-driven instability in a predator-prey system with fear and group defense, Mathematics, 8 (2020), 1–20. https://doi.org/10.3390/math8081244 doi: 10.3390/math8081244
|
| [29] |
J. L. Chen, X. Q. He, F. D. Chen, The influence of fear effect to a discrete-time predator-prey system with predator has other food resource, Mathematics, 9 (2021), 1–20. https://doi.org/10.3390/math9080865 doi: 10.3390/math9080865
|
| [30] |
H. Y. Chen, C. R. Zhang, Dynamic analysis of a Leslie-Gower-type predator-prey system with the fear effect and ratio-dependent Holling Ⅲ functional response, Nonlinear Anal. Model. Control, 27 (2022), 904–926. https://doi.org/10.15388/namc.2022.27.27932 doi: 10.15388/namc.2022.27.27932
|
| [31] |
W. Cresswell, Predation in bird populations, J. Ornithol., 152 (2011), 251–263. https://doi.org/10.1007/s10336-010-0638-1 doi: 10.1007/s10336-010-0638-1
|
| [32] |
M. Clinchy, M. J. Sheriff, L. Y. Zanette, Predator-induced stress and the ecology of fear, Funct. Ecol., 27 (2013), 56–65. https://doi.org/10.1111/1365-2435.12007 doi: 10.1111/1365-2435.12007
|
| [33] | Z. F. Zhang, T. R. Ding, W. Z. Huang, Z. X. Dong, Qualitative theory of differential equations, Beijing: Science Press, 1981. |
| [34] | C. Castillo-Chavez, H. R. Thieme, Asymptotically autonomous epidemic models, Math. Popul. Dyn. Anal. Hetereogeneity, 1 (1995), 33–50. |
| [35] | L. Perko, Differential equations and dynamical systems, 3 Eds., New York: Springer, 2001. https://doi.org/10.1007/978-1-4613-0003-8 |
| [36] | S. Wiggins, Introduction to applied nonlinear dynamical systems and chaos, 2 Eds., New York: Springer, 2003. https://doi.org/10.1007/b97481 |






Figures(7)

Yalong Xue, Fengde Chen, Xiangdong Xie, Shengjiang Chen. An analysis of a predator-prey model in which fear reduces prey birth and death rates[J]. AIMS Mathematics, 2024, 9(5): 12906-12927. doi: 10.3934/math.2024630







 DownLoad:
DownLoad: