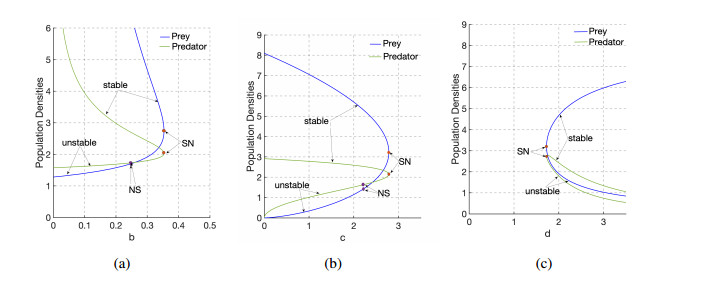
The indirect effect of predation due to fear has proven to have adverse effects on the reproductive rate of the prey population. Here, we present a deterministic two-species predator-prey model with prey herd behavior, mutual interference, and the effect of fear. We give conditions for the existence of some local and global bifurcations at the coexistence equilibrium. We also show that fear can induce extinction of the prey population from a coexistence zone in finite time. Our numerical simulations reveal that varying the strength of fear of predators with suitable choice of parameters can stabilize and destabilize the coexistence equilibrium solutions of the model. Further, we discuss the outcome of introducing a constant harvesting effort to the predator population in terms of changing the dynamics of the system, in particular, from finite time extinction to stable coexistence.
Citation: Kwadwo Antwi-Fordjour, Rana D. Parshad, Hannah E. Thompson, Stephanie B. Westaway. Fear-driven extinction and (de)stabilization in a predator-prey model incorporating prey herd behavior and mutual interference[J]. AIMS Mathematics, 2023, 8(2): 3353-3377. doi: 10.3934/math.2023173
The indirect effect of predation due to fear has proven to have adverse effects on the reproductive rate of the prey population. Here, we present a deterministic two-species predator-prey model with prey herd behavior, mutual interference, and the effect of fear. We give conditions for the existence of some local and global bifurcations at the coexistence equilibrium. We also show that fear can induce extinction of the prey population from a coexistence zone in finite time. Our numerical simulations reveal that varying the strength of fear of predators with suitable choice of parameters can stabilize and destabilize the coexistence equilibrium solutions of the model. Further, we discuss the outcome of introducing a constant harvesting effort to the predator population in terms of changing the dynamics of the system, in particular, from finite time extinction to stable coexistence.

| [1] | A. J. Lotka, Elements of physical biology, Williams and Wilkins, Baltimore. Reprinted as Elements of mathematical biology, Dover, New York, 1925. |
| [2] | V. Volterra, Variazioni e fluttuazioni del numero d'individui in specie animali conviventi, Mem. R. Accad. Naz. Lincei, 2 (1926), 31–113. |
| [3] | G. F. Gause, The struggle for existence, Williams & Wilkins, Baltimore, Maryland, USA, 1934. |
| [4] |
J. R. Beddington, Mutual interference between parasites or predators and its effect on searching efficiency, J. Animal Ecol., 44 (1975), 331–340. https://doi.org/10.2307/3866 doi: 10.2307/3866

|
| [5] |
D. L. DeAngelis, R. A. Goldstein, R. V. ONeill, A model for trophic interaction, Ecology, 56 (1975), 881–892. https://doi.org/10.2307/1936298 doi: 10.2307/1936298
|
| [6] |
C. S. Holling, The components of predation as revealed by a study of small mammal predation of the European pine sawfly, Canad. Entomol., 91 (1959), 293–320. https://doi.org/10.4039/Ent91293-5 doi: 10.4039/Ent91293-5
|
| [7] |
P. A. Abrams, Why ratio dependence is (still) a dad model of predation, Biol. Rev., 90 (2015), 794–814. https://doi.org/10.1111/brv.12134 doi: 10.1111/brv.12134
|
| [8] |
M. A. Aziz-Alaoui, The study of a Leslie-Gower type tri-tropic population models, Chaos, Solitons & Fractals, 14 (2002), 1275–1293. https://doi.org/10.1016/S0960-0779(02)00079-6 doi: 10.1016/S0960-0779(02)00079-6
|
| [9] |
C. S. Holling, The functional response of predators to prey density and its role on mimicry and population regulations, Memoirs of the Entomological Society of Canada, 97 (1965), 5–60. https://doi.org/10.4039/entm9745fv doi: 10.4039/entm9745fv
|
| [10] |
K. Antwi-Fordjour, R. D. Parshad, M. A. Beauregard, Dynamics of a predator-prey model with generalized functional response and mutual interference, Math. Biosci., 360 (2020), 108407. https://doi.org/10.1016/j.mbs.2020.108407 doi: 10.1016/j.mbs.2020.108407
|
| [11] |
R. K. Upadhyay, V. Rai, Why chaos is rarely observed in natural populations, Chaos, Solitons & Fractals, 8 (1997), 1933–1939. https://doi.org/10.1016/S0960-0779(97)00076-3 doi: 10.1016/S0960-0779(97)00076-3
|
| [12] |
A. V. Banerjee, A simple model of herd behavior, Q. J. Econ., 107 (1992), 797–817. https://doi.org/10.2307/2118364 doi: 10.2307/2118364
|
| [13] |
L. Rook, An Economic Psychological Approach to Herd Behavior, J. Econ. Issues, 40 (2006), 75–95. https://doi.org/10.1080/00213624.2006.11506883 doi: 10.1080/00213624.2006.11506883
|
| [14] |
R. M. Raafat, N. Chater, C. Frith, Herding in Humans, Trends Cogn. Sci., 13 (2009), 420–428. https://doi.org/10.1016/j.tics.2009.08.002 doi: 10.1016/j.tics.2009.08.002
|
| [15] | T. B. Veblen, The Theory of the Leisure Class, New York: Dover, 1899. |
| [16] | C. W. Cobb, P. H. Douglas, A theory of production, The American Economic Review, 18 (1928), 139–165. |
| [17] |
V. Ajraldi, M. Pittavino, E. Venturino, Modeling herd behavior in population systems, Nonlinear Anal. Real World Appl., 12 (2011), 2319–2338. https://doi.org/10.1016/j.nonrwa.2011.02.002 doi: 10.1016/j.nonrwa.2011.02.002
|
| [18] |
K. Vilches, E. Gonzalez-Olivares, A Rojas-Palma, Prey herd behavior by a generic non-differentiable functional response, Math. Model. Nat. Phenom., 13 (2018), 26. https://doi.org/10.1051/mmnp/2018038 doi: 10.1051/mmnp/2018038
|
| [19] |
M. L. Rosenzweig, Paradox of enrichment: Destabilization of exploitation ecosystem in ecological time, Science, 171 (1971), 385–387. https://doi.org/10.1126/science.171.3969.385 doi: 10.1126/science.171.3969.385
|
| [20] |
P. A. Braza, Predator–prey dynamics with square root functional responses, Nonlinear Anal.-Real, 13 (2012), 1837–1843. https://doi.org/10.1016/j.nonrwa.2011.12.014 doi: 10.1016/j.nonrwa.2011.12.014
|
| [21] |
B. W. Kooi, E. Venturino, Ecoepidemic predator-prey model with feeding satiation, prey heard behavior and abandoned infected prey, Math. Biosci., 274 (2016), 58–72. https://doi.org/10.1016/j.mbs.2016.02.003 doi: 10.1016/j.mbs.2016.02.003
|
| [22] |
R. D. Parshad, K. Antwi-Fordjour, M. E. Takyi, Some Novel results in two species competition, SIAM J. Appl. Math., 81 (2021), 1847–1869. https://doi.org/10.1137/20M1387274 doi: 10.1137/20M1387274
|
| [23] |
A. Ardito, P Ricciardi, Lyapunov functions for a generalized Gauss-type model, J. Math. Biol., 33 (1995), 816–828. https://doi.org/10.1007/BF00187283 doi: 10.1007/BF00187283
|
| [24] |
E. Saez, I Szanto, A polycycle and limit cycles in a non-differentiable predator-prey model, Proc. Indian Acad. Sci. (Math. Sci.), 117 (2007), 219–231. https://doi.org/10.1007/s12044-007-0018-9 doi: 10.1007/s12044-007-0018-9
|
| [25] |
N. Beroual, T. Sari, A predator-prey system with Holling-type functional response, P. Am. Math. Soc., 148 (2020), 5127–5140. https://doi.org/10.1090/proc/15166 doi: 10.1090/proc/15166
|
| [26] |
A. P. Farrell, J. P. Collins, A. L. Greer, H. R. Thieme, Do fatal infectious diseases eradicate host species? J. Math. Bio., 77 (2018), 2103–2164. https://doi.org/10.1007/s00285-018-1249-3 doi: 10.1007/s00285-018-1249-3
|
| [27] |
H. I. Freedman, Stability analysis of a predator-prey system with mutual interference and density-dependent death rates, B. Math. Biol., 41 (1979), 67–78. https://doi.org/10.1016/S0092-8240(79)80054-3 doi: 10.1016/S0092-8240(79)80054-3
|
| [28] |
M. P. Hassell, Mutual interference between searching insect parasites, J. Anim. Ecol., 40 (1971), 473–486. https://doi.org/10.2307/3256 doi: 10.2307/3256
|
| [29] |
M. P. Hassell, Density dependence in single species population, J. Anim. Ecol., 44 (1975), 283–295. https://doi.org/10.2307/3863 doi: 10.2307/3863
|
| [30] |
M. P. Hassell, G. C. Varley, New inductive population model for insect parasites and its bearing on biological control, Nature, Lond., 223 (1969), 1133–1137. https://doi.org/10.1038/2231133a0 doi: 10.1038/2231133a0
|
| [31] |
R. Arditi, J. M. Callois, Y. Tyutyunov, C. Jost, Does mutual interference always stabilize predator–prey dynamics? A comparison of models, CR Biol., 327 (2004), 1037–1057. https://doi.org/10.1016/j.crvi.2004.06.007 doi: 10.1016/j.crvi.2004.06.007
|
| [32] |
H. I. Freedman, G. S. K. Wolkowicz, Predator-prey systems with group defence: The paradox of enrichment revisited, B. Math. Biol., 48 (1986), 493–508. https://doi.org/10.1016/S0092-8240(86)90004-2 doi: 10.1016/S0092-8240(86)90004-2
|
| [33] |
L. H. Erbe, H. I. Freedman, Modeling persistence and mutual interference among subpopulations of ecological communities, B. Math. Biol., 47 (1985), 295–304. https://doi.org/10.1016/S0092-8240(85)90055-2 doi: 10.1016/S0092-8240(85)90055-2
|
| [34] |
K. Wang, Y. Zhu, Periodic solutions, permanence and global attractivity of a delayed impulsive prey–predator system with mutual interference, Nonlinear Anal.-Real, 14 (2013), 1044–1054. https://doi.org/10.1016/j.nonrwa.2012.08.016 doi: 10.1016/j.nonrwa.2012.08.016
|
| [35] |
R. K. Upadhyay, R. D. Parshad, K. Antwi-Fordjour, E. Quansah, S. Kumari, Global dynamics of stochastic predator-prey with mutual interference and prey defense, J. Appl. Math. Comput., 60 (2019), 169–190. https://doi.org/10.1007/s12190-018-1207-7 doi: 10.1007/s12190-018-1207-7
|
| [36] |
X. Lin, F. D. Chen, Almost periodic solution for a Volterra model with mutual interference and Beddington–DeAngelis functional response, Appl. Math. Comput., 214 (2009), 548–556. https://doi.org/10.1016/j.amc.2009.04.028 doi: 10.1016/j.amc.2009.04.028
|
| [37] |
K. Wang, Existence and global asymptotic stability of positive periodic solution for a predator–prey system with mutual interference, Nonlinear Anal.-Real, 12 (2009), 2774–2783. https://doi.org/10.1016/j.nonrwa.2008.08.015 doi: 10.1016/j.nonrwa.2008.08.015
|
| [38] |
Z. Ma, F. Chen, C Wu, W Chen, Dynamic behaviors of a Lotka–Volterra predator–prey model incorporating a prey refuge and predator mutual interference, Appl. Math. Comput., 219 (2013), 7945–7953. https://doi.org/10.1016/j.amc.2013.02.033 doi: 10.1016/j.amc.2013.02.033
|
| [39] |
L. Zanette, L., A. F. White, M. C. Allen, M. Clinchy, Perceived predation risk reduces the number of offspring songbirds produce per year, Science, 334 (2011), 1398–1401. https://doi.org/10.1126/science.1210908 doi: 10.1126/science.1210908
|
| [40] |
J. P. Suraci, M. Clinchy, L. M. Dill, D. Roberts, L. Y. Zanette, Fear of large carnivores causes a trophic cascade, Nat. Commun., 7 (2016), 10698. https://doi.org/10.1038/ncomms10698 doi: 10.1038/ncomms10698
|
| [41] |
F. Hua, K. E. Sieving, R. J. Fletcher, C. A. Wright, Increased perception of predation risk to adults and offspring alters avian reproductive strategy and performance, Behav. Ecol., 25 (2014), 509–519. https://doi.org/10.1093/beheco/aru017 doi: 10.1093/beheco/aru017
|
| [42] |
A. J. Wirsing, W. J. Ripple, A comparison of shark and wolf research reveals similar behavioral responses by prey, Front. Ecol. Environ., 9 (2011), 335–341. https://doi.org/10.1890/090226 doi: 10.1890/090226
|
| [43] |
A. G. Bauman, J. C. L. Seah, F. A. Januchowski-Hartley, J. Fong, P. A. Todd, Fear effects associated with predator presence and habitat structure interact to alter herbivory on coral reefs, Biol. Lett., 15 (2019), 20190409. https://doi.org/10.1098/rsbl.2019.0409 doi: 10.1098/rsbl.2019.0409
|
| [44] |
X. Wang, L. Zanette, X. Zou, Modelling the fear effect in predator-prey interactions, J. Math. Bio., 73 (2016), 1179–1204. https://doi.org/10.1007/s00285-016-0989-1 doi: 10.1007/s00285-016-0989-1
|
| [45] |
S. Pal, N. Pal, S. Samanta, J. Chattopadhyay, Effect of hunting cooperation and fear in a predator-prey model, Ecol. Complex., 39 (2019), 100770. https://doi.org/10.1016/j.ecocom.2019.100770 doi: 10.1016/j.ecocom.2019.100770
|
| [46] |
R. K. Upadhyay, S. Mishra, Population dynamic consequences of fearful prey in a spatiotemporal predator-prey system, Math. Biosci. Eng., 16 (2019), 338–372. https://doi.org/10.3934/mbe.2019017 doi: 10.3934/mbe.2019017
|
| [47] |
S. Pal, S. Majhi, S. Mandal, N. Pal, Role of Fear in a Predator–Prey Model with Beddington–DeAngelis Functional Response, Z. Naturforsch., 74 (2019), 581–595. https://doi.org/10.1515/zna-2018-0449 doi: 10.1515/zna-2018-0449
|
| [48] |
H. Zhang, Y. Cai, S. Fu, W. Wang, Impact of the fear effect in a prey-predator model incorporating a prey refuge, Appl. Math. Comput., 36 (2019), 328–337. https://doi.org/10.1016/j.amc.2019.03.034 doi: 10.1016/j.amc.2019.03.034
|
| [49] | K. Seonguk, K Antwi-Fordjour, Prey group defense to predator aggregated induced fear, Eur. Phys. J. Plus, 137 (2022), 1–17. |
| [50] |
H. Verma, K. Antwi-Fordjour, M. Hossain, N Pal, R. D. Parshad, P. Mathur, A "Double" Fear Effect in a Tri-tropic Food Chain Model, Eur. Phys. J. Plus, 136 (2021), 1–17. https://doi.org/10.1140/epjp/s13360-021-01900-3 doi: 10.1140/epjp/s13360-021-01900-3
|
| [51] | Z. Xiao, Z. Li, Stability Analysis of a Mutual Interference Predator-prey Model with the Fear Effect, J. Appl. Sci. Eng., 22 (2019), 205–211. |
| [52] |
F. Brauer, A. C. Soudack, Stability regions and transition phenomena for harvested predator–prey systems, J. Math. Biol., 7 (1979), 319–337. https://doi.org/10.1007/BF00275152 doi: 10.1007/BF00275152
|
| [53] |
T. K. Kar, Modelling and analysis of a harvested prey–predator system incorporating a prey refuge, J. Comput. Appl. Math., 185 (2006), 19–33. https://doi.org/10.1016/j.cam.2005.01.035 doi: 10.1016/j.cam.2005.01.035
|
| [54] |
D. Xiao, W. Li, M Han, Dynamics of a ratio-dependent predator–prey model with predator harvesting, J. Math. Anal. Appl., 324 (2006), 14–29. https://doi.org/10.1016/j.jmaa.2005.11.048 doi: 10.1016/j.jmaa.2005.11.048
|
| [55] |
G. Dai, M. Tang, Coexistence region and global dynamics of a harvested predator–prey system, SIAM J. Appl. Math., 58 (1998), 193–210. https://doi.org/10.1137/S0036139994275799 doi: 10.1137/S0036139994275799
|
| [56] |
J. Liu, L. Zhang, Bifurcation analysis in a prey–predator model with nonlinear predator harvesting, J. Franklin I., 353 (2016), 4701–4714. https://doi.org/10.1016/j.jfranklin.2016.09.005 doi: 10.1016/j.jfranklin.2016.09.005
|
| [57] |
H. Fattahpour, W. Nagata, H. R. Z. Zangeneh, Prey–predator dynamics with two predator types and Michaelis–Menten predator harvesting, Differ. Equ. Dyn. Syst., (2019), 1–26. https://doi.org/10.1007/s12591-019-00500-z doi: 10.1007/s12591-019-00500-z
|
| [58] |
S. Chakraborty, S. Pal, N. Bairagi, Predator-prey interaction with harvesting: mathematical study with biological ramifications, Appl. Math. Model., 36 (2011), 4044–4059. https://doi.org/10.1016/j.apm.2011.11.029 doi: 10.1016/j.apm.2011.11.029
|
| [59] |
X. Gao, S. Ishag, S. Fu, W. Li, W. Wang, Bifurcation and Turing pattern formation in a diffusive ratio-dependent predator–prey model with predator harvesting, Nonlinear Anal.-Real, 51 (2020), 102962. https://doi.org/10.1016/j.nonrwa.2019.102962 doi: 10.1016/j.nonrwa.2019.102962
|
| [60] |
N. H. Fakhry, R. K. Naji, The Dynamics of A Square Root Prey-Predator Model with Fear, Iraqi J. Sci., (2020), 139–146. https://doi.org/10.24996/ijs.2020.61.1.15 doi: 10.24996/ijs.2020.61.1.15
|
| [61] | Y. Huang, Z. Li, The Stability of a Predator-Prey Model with Fear Effect in Prey and Square Root Functional Responses, Ann. of Appl. Math., 36 (2020), 186–194. |
| [62] |
D. Sen, S. Ghorai, S. Sharma, M Banerjee, Allee effect in prey's growth reduces the dynamical complexity in prey-predator model with generalist predator, Appl. Math. Model., 91 (2021), 768–790. https://doi.org/10.1016/j.apm.2020.09.046 doi: 10.1016/j.apm.2020.09.046
|
| [63] | L. Perko, Differential equations and dynamical systems, Vol. 7, Springer Science & Business Media, 2013. |
| [64] | J. D. Murray, Mathematical biology, Springer, New York, 1993. |
| [65] |
A. Dhooge, W. Govaerts, Y. A Kuznetsov, H. G. E. Meijer, B. Sautois, New features of the software MatCont for bifurcation analysis of dynamical systems, Math. Comp. Model. Dyn, 14 (2009), 147–175. https://doi.org/10.1080/13873950701742754 doi: 10.1080/13873950701742754
|
| [66] |
P. Panday, N. Pal, S. Samanta, J Chattopadhyay, Stability and bifurcation analysis of a three-species food chain model with fear, Int. J. Bifurcat. Chaos, 28 (2018), 1850009. https://doi.org/10.1142/S0218127418500098 doi: 10.1142/S0218127418500098
|
| [67] |
J. Lyu, P. J. Schofield, K. M. Reaver, M. Beauregard, R. D. Parshad, A comparison of the Trojan Y Chromosome strategy to harvesting models for eradication of nonnative species, Nat. Resour. Model., 33 (2020), e12252. https://doi.org/10.1111/nrm.12252 doi: 10.1111/nrm.12252
|
| [68] |
J. Sugie, R. Kohno, R. Miyazaki, On a predator-prey system of Holling type, P. Am. Math. Soc., 125 (1997), 2041–2050. https://doi.org/10.1090/S0002-9939-97-03901-4 doi: 10.1090/S0002-9939-97-03901-4
|
| [69] |
R. D. Parshad, S. Wickramsooriya, S. Bailey, A remark on "Biological control through provision of additional food to predators: A theoretical study"[Theor. Popul. Biol. 72 (2007) 111–120], Theor. Popul. Biol., 132 (2020), 60–68. https://doi.org/10.1016/j.tpb.2019.11.010 doi: 10.1016/j.tpb.2019.11.010
|
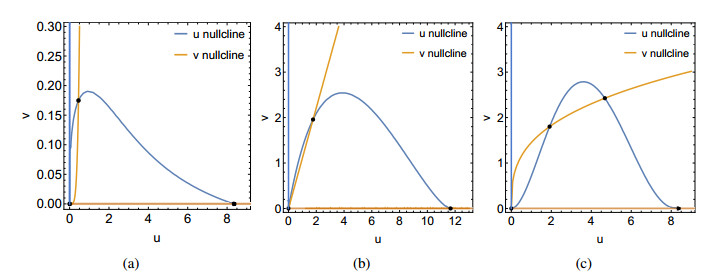
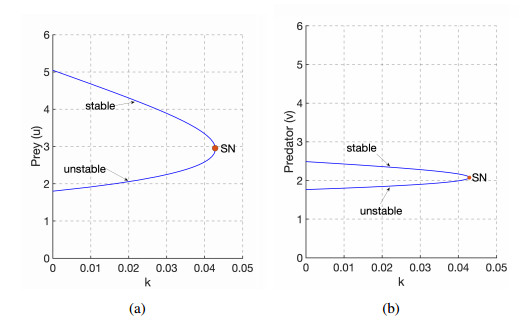
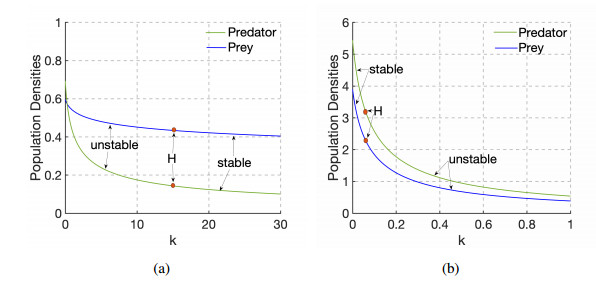
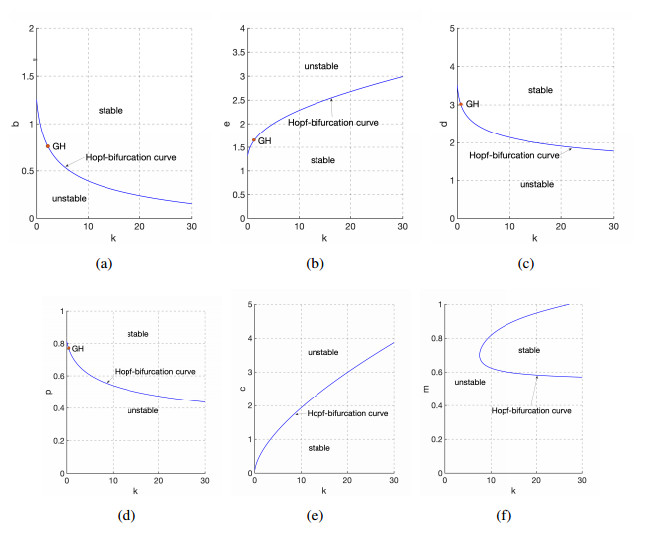
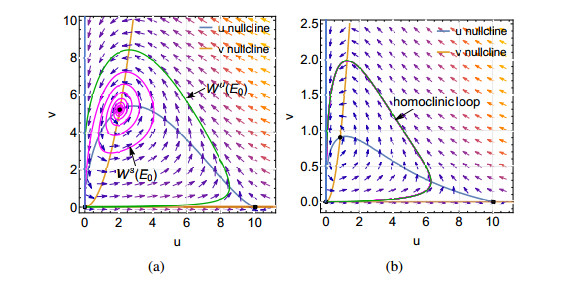
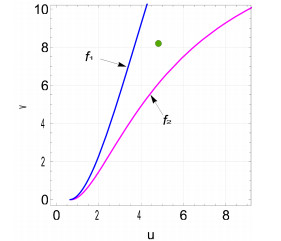
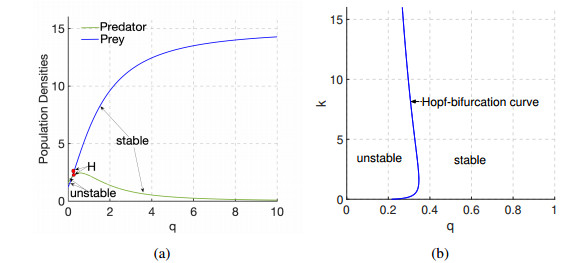
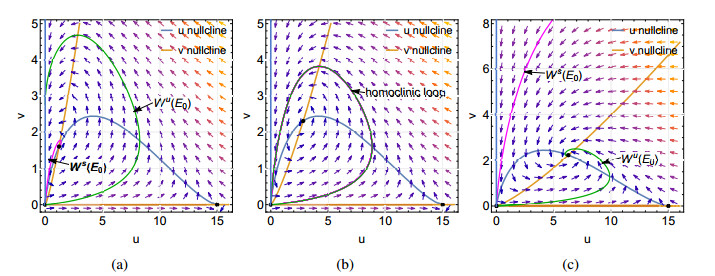
Figures(9)

Kwadwo Antwi-Fordjour, Rana D. Parshad, Hannah E. Thompson, Stephanie B. Westaway. Fear-driven extinction and (de)stabilization in a predator-prey model incorporating prey herd behavior and mutual interference[J]. AIMS Mathematics, 2023, 8(2): 3353-3377. doi: 10.3934/math.2023173







 DownLoad:
DownLoad: