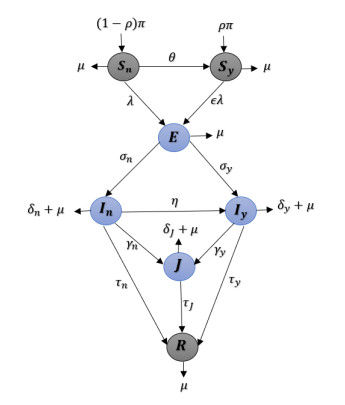
Monkeypox (mpox), a zoonotic viral disease caused by the monkeypox virus (mpoxv), is endemic in many countries in West Africa and is sometimes exported to other parts of the world. The recent outbreak of mpoxv in humans, in endemic and non-endemic countries, has created substantial public health concern worldwide. This research uses a mechanistic model to study the transmission dynamics of mpoxv epidemics in the USA. Our model describes the interaction between different categories of individuals represent various infection phases and hospitalization processes. The model also takes into account the extent of compliance with non-pharmaceutical intervention strategies (NPIs), such as using condoms during sexual contact, quarantine and avoiding large gatherings. The model's equilibria are analyzed, and results on asymptotic stability are obtained. Moreover, the basic reproductive number and other threshold quantities are used to establish the conditions for a forward or backward bifurcation. Our model accurately captures the incidence curves from mpox surveillance data for the USA, indicating that it can be used to explain mpoxv transmission and suggest some effective ways to enhance control efforts. In addition, numerical simulations are carried out to examine the influence of some parameters on the overall dynamics of the model. A partial rank correlation coefficient is adopted for the sensitivity analysis to determine the model most important parameters, which require close attention for effective mpoxv prevention and control. We conclude that it is especially important to ensure that NPIs are properly followed to mitigate mpoxv outbreaks effectively.
Citation: Rubayyi T. Alqahtani, Salihu S. Musa, Mustafa Inc. Modeling the role of public health intervention measures in halting the transmission of monkeypox virus[J]. AIMS Mathematics, 2023, 8(6): 14142-14166. doi: 10.3934/math.2023723
Monkeypox (mpox), a zoonotic viral disease caused by the monkeypox virus (mpoxv), is endemic in many countries in West Africa and is sometimes exported to other parts of the world. The recent outbreak of mpoxv in humans, in endemic and non-endemic countries, has created substantial public health concern worldwide. This research uses a mechanistic model to study the transmission dynamics of mpoxv epidemics in the USA. Our model describes the interaction between different categories of individuals represent various infection phases and hospitalization processes. The model also takes into account the extent of compliance with non-pharmaceutical intervention strategies (NPIs), such as using condoms during sexual contact, quarantine and avoiding large gatherings. The model's equilibria are analyzed, and results on asymptotic stability are obtained. Moreover, the basic reproductive number and other threshold quantities are used to establish the conditions for a forward or backward bifurcation. Our model accurately captures the incidence curves from mpox surveillance data for the USA, indicating that it can be used to explain mpoxv transmission and suggest some effective ways to enhance control efforts. In addition, numerical simulations are carried out to examine the influence of some parameters on the overall dynamics of the model. A partial rank correlation coefficient is adopted for the sensitivity analysis to determine the model most important parameters, which require close attention for effective mpoxv prevention and control. We conclude that it is especially important to ensure that NPIs are properly followed to mitigate mpoxv outbreaks effectively.

| [1] |
N. Ahmed, A. Elsonbaty, A. Raza, M. Rafiq, W. Adel, Numerical simulation and stability analysis of a novel reaction-diffusion COVID-19 model, Nonlinear Dyn., 106 (2021), 1293–1310. http://dx.doi.org/10.1007/s11071-021-06623-9 doi: 10.1007/s11071-021-06623-9

|
| [2] |
N. Ahmed, M. Rafiq, W. Adel, H. Rezazadeh, I. Khan, K. Nisar, Structure preserving numerical analysis of HIV and CD4+ T-cells reaction diffusion model in two space dimensions, Chaos Soliton. Fract., 139 (2020), 110307. http://dx.doi.org/10.1016/j.chaos.2020.110307 doi: 10.1016/j.chaos.2020.110307
|
| [3] |
J. Americo, P. Earl, B. Moss, Virulence differences of mpox (monkeypox) virus clades Ⅰ, Ⅱa, and Ⅱb. 1 in a small animal model, PNAS, 120 (2023), 2220415120. http://dx.doi.org/10.1073/pnas.2220415120 doi: 10.1073/pnas.2220415120
|
| [4] |
S. Bankuru, S. Kossol, W. Hou, P. Mahmoudi, J. Rychtář, D. Taylor, A game-theoretic model of Monkeypox to assess vaccination strategies, PeerJ, 8 (2020), 9272. http://dx.doi.org/10.7717/peerj.9272 doi: 10.7717/peerj.9272
|
| [5] | J. Carr, Applications of centre manifold theory, New York: Springer, 1981. http://dx.doi.org/10.1007/978-1-4612-5929-9 |
| [6] |
C. Castillo-Chavez, B. Song, Dynamical models of tuberculosis and their applications, Math. Biosci. Eng., 1 (2004), 361–404. http://dx.doi.org/10.3934/mbe.2004.1.361 doi: 10.3934/mbe.2004.1.361
|
| [7] | 2022 Mpox outbreak global map, Centers for Disease Control and Prevention, 2023. Available from: https://www.cdc.gov/poxvirus/monkeypox/response/2022/world-map.html |
| [8] | 2022 outbreak cases and data, Centers for Disease Control and Prevention, 2023. Available from: https://www.cdc.gov/poxvirus/monkeypox/response/2022/index.html |
| [9] | Life expectancy in the US dropped for the second year in a row in 2021, Centers for Disease Control and Prevention, 2022. Available from: https://www.cdc.gov/nchs/pressroom/nchs_press_releases/2022/20220831.htm |
| [10] |
O. Diekmann, J. Heesterbeek, J. Metz, On the definition and the computation of the basic reproduction ratio R0 in models for infectious diseases in heterogeneous populations, J. Math. Biol., 28 (1990), 365–382. http://dx.doi.org/10.1007/BF00178324 doi: 10.1007/BF00178324
|
| [11] | B. Dubey, P. Dubey, U. Dubey, Dynamics of an SIR model with nonlinear incidence and treatment rate, Appl. Appl. Math., 10 (2015), 5. |
| [12] |
A. El-Mesady, A. Elsonbaty, W. Adel, On nonlinear dynamics of a fractional order monkeypox virus model, Chaos Soliton. Fract., 164 (2022), 112716. http://dx.doi.org/10.1016/j.chaos.2022.112716 doi: 10.1016/j.chaos.2022.112716
|
| [13] |
A. Elsonbaty, Z. Sabir, R. Ramaswamy, W. Adel, Dynamical analysis of a novel discrete fractional SITRS model for COVID-19, Fractals, 29 (2021), 2140035. http://dx.doi.org/10.1142/S0218348X21400351 doi: 10.1142/S0218348X21400351
|
| [14] |
A. Endo, H. Murayama, S. Abbott, R. Ratnayake, C. Pearson, W. Edmunds, et al., Heavy-tailed sexual contact networks and monkeypox epidemiology in the global outbreak, Science, 378 (2022), 90–94. http://dx.doi.org/10.1126/science.add4507 doi: 10.1126/science.add4507
|
| [15] |
S. Garba, A. Gumel, M. Bakar, Backward bifurcations in dengue transmission dynamics, Math. Biosci., 215 (2008), 11–25. http://dx.doi.org/10.1016/j.mbs.2008.05.002 doi: 10.1016/j.mbs.2008.05.002
|
| [16] |
D. Gao, Y. Lou, D. He, T. Porco, Y. Kuang, G. Chowell, et al., Prevention and control of Zika as a mosquito-borne and sexually transmitted disease: a mathematical modeling analysis, Sci. Rep., 6 (2016), 28070. http://dx.doi.org/10.1038/srep28070 doi: 10.1038/srep28070
|
| [17] |
I. Ghosh, P. Tiwari, J. Chattopadhyay, Effect of active case finding on dengue control: implications from a mathematical model, J. Theor. Biol., 464 (2019), 50–62. http://dx.doi.org/10.1016/j.jtbi.2018.12.027 doi: 10.1016/j.jtbi.2018.12.027
|
| [18] |
A. Gumel, Causes of backward bifurcations in some epidemiological models, J. Math. Anal. Appl., 395 (2012), 355–365. http://dx.doi.org/10.1016/j.jmaa.2012.04.077 doi: 10.1016/j.jmaa.2012.04.077
|
| [19] | H. Gunerhan, H. Rezazadeh, W. Adel, M. Hatami, K. Sagayam, H. Emadifar, et al., Analytical approximate solution of fractional order smoking epidemic model, Adv. Mech. Eng., in press. http://dx.doi.org/10.1177/16878132221123888 |
| [20] |
H. Günerhan, H. Dutta, M. Dokuyucu, W. Adel, Analysis of a fractional HIV model with Caputo and constant proportional Caputo operators, Chaos Soliton. Fract., 139 (2020), 110053. http://dx.doi.org/10.1016/j.chaos.2020.110053 doi: 10.1016/j.chaos.2020.110053
|
| [21] |
C. Happi, I. Adetifa, P. Mbala, R. Njouom, E. Nakoune, A. Happi, et al., Urgent need for a non-discriminatory and non-stigmatizing nomenclature for monkeypox virus, PLoS Biol., 20 (2022), 3001769. http://dx.doi.org/10.1371/journal.pbio.3001769 doi: 10.1371/journal.pbio.3001769
|
| [22] |
B. Hernaez, A. Muñoz-Gómez, A. Sanchiz, E. Orviz, A. Valls-Carbo, I. Sagastagoitia, et al., Monitoring monkeypox virus in saliva and air samples in Spain: a cross-sectional study, The Lancet Microbe, 4 (2023), 21–28. http://dx.doi.org/10.1016/S2666-5247(22)00291-9 doi: 10.1016/S2666-5247(22)00291-9
|
| [23] |
N. Hussaini, K. Okuneye, A. Gumel, Mathematical analysis of a model for zoonotic visceral leishmaniasis, Infect. Dis. Model., 2 (2017), 455–474. http://dx.doi.org/10.1016/j.idm.2017.12.002 doi: 10.1016/j.idm.2017.12.002
|
| [24] |
M. Izadi, S. Yüzbaşı, W. Adel, Accurate and efficient matrix techniques for solving the fractional Lotka-Volterra population model, Physica A, 600 (2022), 127558. http://dx.doi.org/10.1016/j.physa.2022.127558 doi: 10.1016/j.physa.2022.127558
|
| [25] |
A. Khan, M. Naveed, M. Dur-e-Ahmad, M. Imran, Estimating the basic reproductive ratio for the Ebola outbreak in Liberia and Sierra Leone, Infect. Dis. Poverty, 4 (2015), 13. http://dx.doi.org/10.1186/s40249-015-0043-3 doi: 10.1186/s40249-015-0043-3
|
| [26] |
N. Hussaini, M. Winter, A. Gumel, Qualitative assessment of the role of public health education program on HIV transmission dynamics, Math. Med. Biol., 28 (2011), 245–270. http://dx.doi.org/10.1093/imammb/dqq009 doi: 10.1093/imammb/dqq009
|
| [27] | J. La Salle, The stability of dynamical systems, Philadelphia: SIAM, 1976. |
| [28] |
Q. Lin, S. Musa, S. Zhao, D. He, Modeling the 2014–2015 Ebola virus disease outbreaks in Sierra Leone, Guinea, and Liberia with effect of high-and low-risk susceptible individuals, Bull. Math. Biol., 82 (2020), 102. http://dx.doi.org/10.1007/s11538-020-00779-y doi: 10.1007/s11538-020-00779-y
|
| [29] |
S. Musa, Z. Abdullahi, S. Zhao, U. Bello, N. Hussaini, A. Habib, et al., Transmission dynamics of Monkeypox virus in Nigeria during the current COVID-19 pandemic and estimation of effective reproduction number, Vaccines, 10 (2022), 2153. http://dx.doi.org/10.3390/vaccines10122153 doi: 10.3390/vaccines10122153
|
| [30] |
S. Musa, N. Hussaini, S. Zhao, D. He, Dynamical analysis of chikungunya and dengue co-infection model, Discrete Cont. Dyn.-B, 25 (2020), 1907–1933. http://dx.doi.org/10.3934/dcdsb.2020009 doi: 10.3934/dcdsb.2020009
|
| [31] |
S. Musa, A. Yusuf, E. Bakare, Z. Abdullahi, L. Adamu, U. Mustapha, et al., Unravelling the dynamics of Lassa fever transmission with differential infectivity: Modeling analysis and control strategies, Math. Biosci. Eng., 19 (2022), 13114–13136. http://dx.doi.org/10.3934/mbe.2022613 doi: 10.3934/mbe.2022613
|
| [32] |
S. Musa, S. Zhao, D. Gao, Q. Lin, G. Chowell, D. He, Mechanistic modelling of the large-scale Lassa fever epidemics in Nigeria from 2016 to 2019, J. Theor. Biol., 493 (2020), 110209. http://dx.doi.org/10.1016/j.jtbi.2020.110209 doi: 10.1016/j.jtbi.2020.110209
|
| [33] |
L. Nkamba, T. Manga, F. Agouanet, M. Mann Manyombe, Mathematical model to assess vaccination and effective contact rate impact in the spread of tuberculosis, J. Biol. Dyn., 13 (2019), 26–42. http://dx.doi.org/10.1080/17513758.2018.1563218 doi: 10.1080/17513758.2018.1563218
|
| [34] |
D. Ogoina, M. Iroezindu, H. James, R. Oladokun, A. Yinka-Ogunleye, P. Wakama, et al., Clinical course and outcome of human Monkeypox in Nigeria, Clin. Infect. Dis., 71 (2020), 210–214. http://dx.doi.org/10.1093/cid/ciaa143 doi: 10.1093/cid/ciaa143
|
| [35] |
D. Ogoina, J. Izibewule, A. Ogunleye, E. Ederiane, U. Anebonam, A. Neni, et al., The 2017 human monkeypox outbreak in Nigeria-report of outbreak experience and response in the Niger Delta University Teaching Hospital, Bayelsa State, Nigeria, PLoS. One, 14 (2019), 0214229. http://dx.doi.org/10.1371/journal.pone.0214229 doi: 10.1371/journal.pone.0214229
|
| [36] |
O. Peter, S. Kumar, N. Kumari, F. Oguntolu, K. Oshinubi, R. Musa, Transmission dynamics of Monkeypox virus: a mathematical modelling approach, Model. Earth Syst. Environ., 8 (2022), 3423–3434. http://dx.doi.org/10.1007/s40808-021-01313-2 doi: 10.1007/s40808-021-01313-2
|
| [37] |
O. Peter, F. Oguntolu, M. Ojo, A. Oyeniyi, R. Jan, I. Khan, Fractional order mathematical model of monkeypox transmission dynamics, Phys. Scr., 97 (2022), 084005. http://dx.doi.org/10.1088/1402-4896/ac7ebc doi: 10.1088/1402-4896/ac7ebc
|
| [38] |
J. Riopelle, V. Munster, J. Port, Atypical and unique transmission of monkeypox virus during the 2022 outbreak: an overview of the current state of knowledge, Viruses, 14 (2022), 2012. http://dx.doi.org/10.3390/v14092012 doi: 10.3390/v14092012
|
| [39] |
P. Roop-O, W. Chinviriyasit, S. Chinviriyasit, The effect of incidence function in backward bifurcation for malaria model with temporary immunity, Math. Biosci., 265 (2015), 47–64. http://dx.doi.org/10.1016/j.mbs.2015.04.008 doi: 10.1016/j.mbs.2015.04.008
|
| [40] |
M. Safi, A. Gumel, Qualitative study of a quarantine/isolation model with multiple disease stages, Appl. Math. Comput., 218 (2011), 1941–1961. http://dx.doi.org/10.1016/j.amc.2011.07.007 doi: 10.1016/j.amc.2011.07.007
|
| [41] |
Z. Shuai, P. van den Driessche, Global stability of infectious disease models using Lyapunov functions, SIAM J. Appl. Math., 73 (2013), 1513–1532. http://dx.doi.org/10.1137/120876642 doi: 10.1137/120876642
|
| [42] |
G. Sun, J. Xie, S. Huang, Z. Jin, M. Li, L. Liu, Transmission dynamics of cholera: mathematical modeling and control strategies, Commun. Nonlinear Sci., 45 (2017), 235–244. http://dx.doi.org/10.1016/j.cnsns.2016.10.007 doi: 10.1016/j.cnsns.2016.10.007
|
| [43] |
C. Trotter, N. Gay, W. Edmunds, Dynamic models of meningococcal carriage, disease, and the impact of serogroup C conjugate vaccination. Am. J. Epidemiol., 162 (2005), 89–100. http://dx.doi.org/10.1093/aje/kwi160 doi: 10.1093/aje/kwi160
|
| [44] |
P. van den Driessche, Reproduction numbers of infectious disease models, Infectious Disease Modelling, 2 (2017), 288–303. http://dx.doi.org/10.1016/j.idm.2017.06.002 doi: 10.1016/j.idm.2017.06.002
|
| [45] |
P. van den Driessche, J. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180 (2002), 29–48. http://dx.doi.org/10.1016/S0025-5564(02)00108-6 doi: 10.1016/S0025-5564(02)00108-6
|
| [46] |
T. Ward, R. Christie, R. Paton, F. Cumming, C. Overton, Transmission dynamics of monkeypox in the United Kingdom: contact tracing study, BMJ, 379 (2022), 073153. http://dx.doi.org/10.1136/bmj-2022-073153 doi: 10.1136/bmj-2022-073153
|
| [47] | Monkeypox, World Health Organization, 2022. Available from: https://www.who.int/news-room/fact-sheets/detail/monkeypox?gclid = CjwKCAjws–ZBhAXEiwAv-RNL3T7SFlwhw8Ew8-flekFNnaO9DZaJgz0ybxgg7xwzsIRYi_VAYCb1hoC1OIQAvD_BwE |
| [48] | Population of the United States (2020 and historical), Worldometer, 2022. Available from: https://www.worldometers.info/world-population/us-population/. |
| [49] |
S. Yang, X. Guo, Z. Zhao, B. Abudunaibi, Y. Zhao, J. Rui, et al., Possibility of mpox viral transmission and control from high-risk to the general population: a modeling study, BMC Infect. Dis., 23 (2023), 119. http://dx.doi.org/10.1186/s12879-023-08083-5 doi: 10.1186/s12879-023-08083-5
|
| [50] |
C. Yang, X. Wang, D. Gao, J. Wang, Impact of awareness programs on cholera dynamics: two modeling approaches, Bull. Math. Biol., 79 (2017), 2109–2131. http://dx.doi.org/10.1007/s11538-017-0322-1 doi: 10.1007/s11538-017-0322-1
|
| [51] |
P. Yuan, Y. Tan, L. Yang, E. Aruffo, N. Ogden, J. Bélair, et al., Assessing transmission risks and control strategy for monkeypox as an emerging zoonosis in a metropolitan area, J. Med. Virol., 95 (2023), 28137. http://dx.doi.org/10.1002/jmv.28137 doi: 10.1002/jmv.28137
|
| [52] |
P. Yuan, Y. Tan, L. Yang, E. Aruffo, N. Ogden, J. Belair, et al., Modelling vaccination and control strategies for outbreaks of monkeypox at gatherings, Front. Public Health, 10 (2022), 1026489. http://dx.doi.org/10.3389/fpubh.2022.1026489 doi: 10.3389/fpubh.2022.1026489
|
| [53] |
M. Al-Shomrani, S. Musa, A. Yusuf, Unfolding the transmission dynamics of monkeypox virus: an epidemiological modelling analysis, Mathematics, 11 (2023), 1121. http://dx.doi.org/10.3390/math11051121 doi: 10.3390/math11051121
|
| [54] |
Z. Yuan, S. Musa, S. Hsu, C. Cheung, D. He, Post pandemic fatigue: what are effective strategies? Sci. Rep., 12 (2022), 9706. http://dx.doi.org/10.1038/s41598-022-13597-0 doi: 10.1038/s41598-022-13597-0
|
| [55] |
S. Zhao, L. Stone, D. Gao, D. He, Modelling the large-scale yellow fever outbreak in Luanda, Angola, and the impact of vaccination, PLoS Neglect. Trop. Dis., 12 (2018), 0006158. http://dx.doi.org/10.1371/journal.pntd.0006158 doi: 10.1371/journal.pntd.0006158
|
| [56] |
A. Zumla, S. Valdoleiros, N. Haider, D. Asogun, F. Ntoumi, E. Petersen, et al., Monkeypox outbreaks outside endemic regions: scientific and social priorities, Lancet Infect. Dis., 22 (2022), 929–931. http://dx.doi.org/10.1016/S1473-3099(22)00354-1 doi: 10.1016/S1473-3099(22)00354-1
|





Figures(6) / Tables(2)

Rubayyi T. Alqahtani, Salihu S. Musa, Mustafa Inc. Modeling the role of public health intervention measures in halting the transmission of monkeypox virus[J]. AIMS Mathematics, 2023, 8(6): 14142-14166. doi: 10.3934/math.2023723







 DownLoad:
DownLoad: