Citation: Tailei Zhang, Hui Li, Na Xie, Wenhui Fu, Kai Wang, Xiongjie Ding. Mathematical analysis and simulation of a Hepatitis B model with time delay: A case study for Xinjiang, China[J]. Mathematical Biosciences and Engineering, 2020, 17(2): 1757-1775. doi: 10.3934/mbe.2020092

| [1] | World Health Organization, Hepatitis B, 2018. Available from: https://www.who.int/newsroom/fact-sheets/detail/hepatitis-b. |
| [2] | Y. Cui, M. Moriyama, M. Rahman, Analysis of the incidence of hepatitis B and hepatitis C and association with socio-economic factors in various regions in China, Health, 10 (2018), 1210-1220. |
| [3] | Z. Sun, L. Ming, X. Zhu, J. Lu, Prevention and Control of Hepatitis B in China, J. Med. Virol., 67 (2002), 447-450. |
| [4] | China Center for Disease Control and Prevention, Questions and answers on hepatitis B vaccination, 2013.Available from: http://www.chinacdc.cn/zxdt/201312/t20131230_92034.htm. |
| [5] | M. Nowak, S. Bonhoeffer, A. M. Hill, R. Boehme, H. C. Thomas, H. Mcdade, Viral dynamics in hepatitis B virus infection,Proc. Natl. Acad. Sci. U. S. A., 93 (1996), 4398-4402. |
| [6] | S. M. Ciupe, R. M. Ribeiro, P. W. Nelson, A. S. Perelson, Modeling the mechanisms of acute hepatitis B virus infection,J. Theor. Biol., 247 (2007), 23-35. |
| [7] | R. Xu and Z. Ma, An HBV model with diffusion and time delay, J. Theor. Biol., 257 (2009), 499-509. |
| [8] | P. M. Tchinda, J. J. Tewa, B. Mewoli, S. Bowong, A Theoretical assessment of the effects of distributed delay of the transmission dynamics of hepatitis B,J. Biol. Syst., 23 (2015), 423-455. |
| [9] | K. Hattaf, N. Yousfi, A. Tridane, Stability analysis of a virus dynamics model with general incidence rate and two delays,Appl. Math. Comput., 221 (2013), 514-521. |
| [10] | K. Wang, W. Wang, S. Song,Dynamics of an HBV model with diffusion and delay,J. Theor. Biol., 253 (2008), 36-44. |
| [11] | K. Wang, A. Fan, A. Torres, Global properties of an improved hepatitis B virus model, Nonlinear Anal.-Real World Appl., 11 (2010), 3131-3138. |
| [12] | S. Zhao, Z. Xu, Y. Lu,A mathematical model of hepatitis B virus transmission and its application for vaccination strategy in China,Int. J. Epidemiol., 29 (2000), 744-752. |
| [13] | L. Zou, S. Ruan, W. Zhang, On the sexual transmission dynamics of hepatitis B virus in China, J. Theor. Biol., 369 (2015), 1-12. |
| [14] | S. Zhang, Y. Zhou, Dynamic analysis of a hepatitis B model with three-age-classes, Commun. Nonlinear. Sci., 19 (2014), 2466-2478. |
| [15] | S. Zhang, X. Xu, A mathematical model for hepatitis B with infection-age structure, Discrete Contin. Dyn. Syst.-Ser. B, 21 (2016), 1329-1346. |
| [16] | J. Pang, J. Cui, X. Zhou, Dynamical behavior of a hepatitis B virus transmission model with vaccination,J. Theor. Biol.,265 (2010), 572-578. |
| [17] | L. Zou, S. Ruan, W. Zhang, An age-structured model for the transmission dynamics of hepatitis B,SIAM J. Appl. Math., 70 (2010), 3121-3139. |
| [18] | B. O. Emerenini, S. Inyama, Mathematical model and analysis of hepatitis B virus transmission dynamics,F1000 Research,7 (2017). |
| [19] | R. Akbari, A. V. Kamyad, A. A. Heydari, Stability analysis of the transmission dynamics of an HBV model,Int. J. Indu. Math., 8 (2016), 119-219. |
| [20] | J. Mann, M. Roberts, Modelling the epidemiology of hepatitis B in New Zealand, J. Theor. Biol., 269 (2011), 266-272. |
| [21] | World Health Organization, Media centre, 2014. Available from: http://www.who.int/mediacentre/factsheets/fs204/en/. |
| [22] | X. Q. Zhao,Basic reproduction ratios for periodic compartmental models with time delay,J. Dyn. Differ. Equ., 29 (2017), 67-82. |
| [23] | Y. Kuang, Delay differential equations with application to population dynamics, Academic Press, San Diego, 1993. |
| [24] | C. C. McCluskey, Complete global stability for an SIR epidemic model with delay-Distributed or discrete,Nonlinear Anal. RWA 11 (2010), 55-59. |
| [25] | C. Vargas-De-León, Stability analysis of a model for HBV infection with cure of infected cells and intracellular delay,Appl. Math. Comput., 219 (2012), 389-398. |
| [26] | X. Q. Zhao,Dynamical systems in population biology,Springer, New York, 2017. |
| [27] | T. Zhang, K. Wang, X. Zhang, Modeling and analyzing the transmission dynamics of HBV Epidemic in Xinjiang, China,PLoS ONE, 10 (2015), e0138765. |
| [28] | S. Zhang and Y. Zhou, The analysis and application of an HBV model, Appl. Math. Model., 36 (2012), 1302-1312. |
| [29] | L. Zou, W. Zhang, S. Ruan, Modeling the transmission dynamics and control of hepatitis B virus in China, J. Theor. Biol., 262 (2010), 330-338. |
| [30] |
P. Mathurin, C. Mouquet, T. Poynard, C. Sylla, H. Benalia, C. Fretz, et al., Impact of hepatitis B and C virus on kidney transplantation outcome,Hepatology, 29 (1999), 257-263. doi: 10.1002/hep.510290123

|
| [31] | S. Marino, I. B. Hogue, C. J. Ray, D. E. Kirschner, A methodology for performing global uncertainty and sensitivity analysis in systems biology,J. Theor. Biol., 254 (2008), 178-196. |
| [32] | World Health Organization,Global hepatitis report, 2017. |
| [33] | J. K. Hale,Theory of functional differential equations,Springer-Verlag, New York, 1977. |
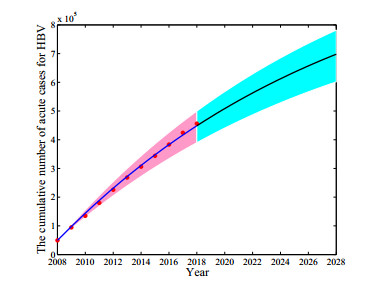
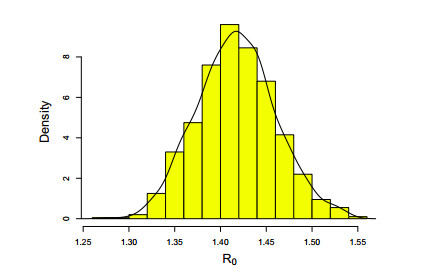
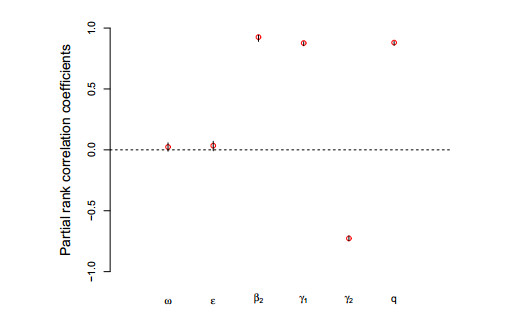
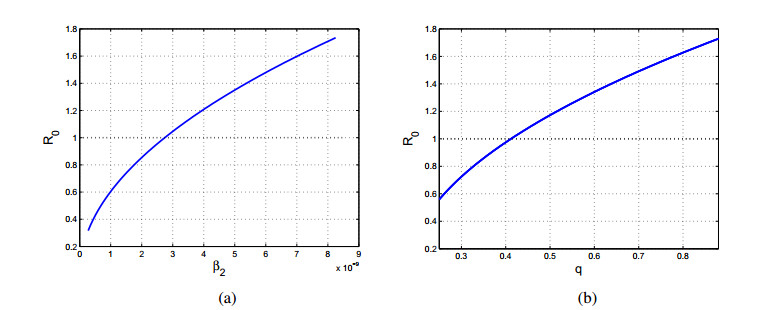
Figures(8) / Tables(3)

Tailei Zhang, Hui Li, Na Xie, Wenhui Fu, Kai Wang, Xiongjie Ding. Mathematical analysis and simulation of a Hepatitis B model with time delay: A case study for Xinjiang, China[J]. Mathematical Biosciences and Engineering, 2020, 17(2): 1757-1775. doi: 10.3934/mbe.2020092







 DownLoad:
DownLoad: