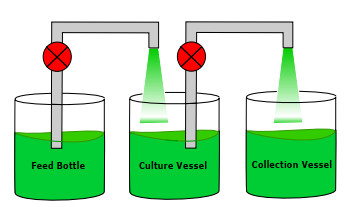
In this paper we study some chemostat models with random bounded fluctuations on the input flow. We start with the classical chemostat system and obtain new models incorporating, for instance, wall growth and different consumption functions, motivated by phenomena in real devices. In every case, we prove existence and uniqueness of positive global solution, existence of deterministic absorbing and attracting sets and we investigate the internal structure of the attracting sets to obtain detailed information about the long-time dynamics of the systems. This allows us to provide conditions under which either extinction or persistence of the species is ensured, the main goal for practitioners. In addition, we provide several numerical simulations to support the theoretical results.
Citation: Tomás Caraballo, Javier López-de-la-Cruz. Survey on chemostat models with bounded random input flow[J]. Mathematical Modelling and Control, 2021, 1(1): 52-78. doi: 10.3934/mmc.2021005
In this paper we study some chemostat models with random bounded fluctuations on the input flow. We start with the classical chemostat system and obtain new models incorporating, for instance, wall growth and different consumption functions, motivated by phenomena in real devices. In every case, we prove existence and uniqueness of positive global solution, existence of deterministic absorbing and attracting sets and we investigate the internal structure of the attracting sets to obtain detailed information about the long-time dynamics of the systems. This allows us to provide conditions under which either extinction or persistence of the species is ensured, the main goal for practitioners. In addition, we provide several numerical simulations to support the theoretical results.

| [1] | J. Andrews, A mathematical model for the continuous culture of microorganisms utilizing inhibitory substrates, Biotechnology & Bioengineering, 10 (1968), 707–723. |
| [2] | L. Arnold, Random Dynamical Systems, Springer Berlin Heidelberg, 1998. |
| [3] | J. Barlow, F. de Noyelles, B. Peterson, J. Peterson and W. Schaffner, Continuous flow nutrient bioassays with natural phytoplankton populations, G. Glass (Editor): Bioassay Techniques and Environmental Chemistry, John Wiley & Sons Ltd., 1973. |
| [4] |
H. R. Bungay and M. L. Bungay, Microbial interactions in continuous culture, Advances in Applied Microbiology, 10 (1968), 269–290. doi: 10.1016/S0065-2164(08)70194-1

|
| [5] |
T. Caraballo, R. Colucci, J. López-de-la-Cruz and A. Rapaport, A way to model stochastic perturbations in population dynamics models with bounded realizations, Communications in Nonlinear Science and Numerical Simulation, 77 (2019), 239–257. doi: 10.1016/j.cnsns.2019.04.019
|
| [6] |
T. Caraballo, R. Colucci, J. López-de-la-Cruz and A. Rapaport, Study of the chemostat model with non-monotonic growth under random disturbances on the removal rate, Mathematical Biosciences and Engineering, 17 (2020), 7480–7501. doi: 10.3934/mbe.2020382
|
| [7] | T. Caraballo, M. J. Garrido-Atienza, J. López-de-la-Cruz and A. Rapaport, Modeling and analysis of random and stochastic input flows in the chemostat model, Discrete & Continuous Dynamical Systems - Series B, 24 (2018), 3591–3614. |
| [8] | T. Caraballo and X. Han, Applied Nonautonomous and Random Dynamical Systems, Applied Dynamical Systems, Springer International Publishing, 2016. |
| [9] |
T. Caraballo, P. E. Kloeden and B. Schmalfuss, Exponentially stable stationary solutions for stochastic evolution equations and their perturbation, Applied Mathematics and Optimization, 50 (2004), 183–207. doi: 10.1007/s00245-004-0802-1
|
| [10] |
T. Caraballo, R. Colucci, J. López-de-la-Cruz and A. Rapaport, Study of the chemostat model with non-monotonic growth under random disturbances on the removal rate, AIMS Mathematical Biosciences and Engineering, 17 (2020), 7480–7501. doi: 10.3934/mbe.2020382
|
| [11] | T. Caraballo, M. J. Garrido-Atienza and J. López-de-la-Cruz, Some Aspects Concerning the Dynamics of Stochastic Chemostats, vol. 69, chapter 11,227–246, Springer International Publishing, 2016. |
| [12] | T. Caraballo, J.López-de-la-Cruz and A. Rapaport, Modeling bounded random fluctuations in biological systems: application to the chemostat model with two species, IFAC-PapersOnLine, 52 (2019), 187–192. |
| [13] |
T. Caraballo and J. López-de-la-Cruz, Bounded random fluctuations on the input flow in chemostat models with wall growth and non-monotonic kinetics, AIMS Mathematics, 6 (2021), 4025–4052. doi: 10.3934/math.2021239
|
| [14] |
I. F. Creed, D. M. McKnight, B. A. Pellerin, M. B. Green, B. A. Bergamaschi, G. R. Aiken et al., The river as a chemostat: fresh perspectives on dissolved organic matter flowing down the river continuum, Canadian Journal of Fisheries and Aquatic Sciences, 72 (2015), 1272–1285. doi: 10.1139/cjfas-2014-0400
|
| [15] | A. Cunningham and R. M. Nisbet, Transients and oscillations in continuous cultures, Mathematics in Microbiology, 77–103. |
| [16] | G. D'Ans, P. Kokotovic and D. Gottlieb, A nonlinear regulator problem for a model of biological waste treatment, IEEE Transactions on Automatic Control, 16 (1971), 341–347. |
| [17] | A. Fredrickson and G. Stephanopoulos, Microbial competition, Science, 213 (1981), 972–979. |
| [18] |
H. I. Freedman and P. Moson, Persistence definitions and their connections, Proceedings of the American Mathematical Society, 109 (1990), 1025–1033. doi: 10.1090/S0002-9939-1990-1012928-6
|
| [19] | R. Freter, Human Intestinal Microflora in Health and Disease, chapter Mechanisms that control the microflora in the large intestine, 33–54, Academic Press, New York, 1983. |
| [20] | R. Freter, An understanding of colonization of the large intestine requires mathematical analysis, Microecology and Therapy, 16 (1986), 147–155. |
| [21] | J. S. H. Haldane, Enzymes, Longmans Green and Co, London, 1930. |
| [22] | J. Harmand, C. Lobry, A. Rapaport and T. Sari, The Chemostat: Mathematical Theory of Micro-organisms Cultures, Wiley, Chemical Engineering Series, John Wiley & Sons, Inc., 2017. |
| [23] |
H. W. Jannasch, Steady state and the chemostat in ecology, Limnology and Oceanography, 19 (1974), 716–720. doi: 10.4319/lo.1974.19.4.0716
|
| [24] |
J. Kalff and R. Knoechel, Phytoplankton and their dynamics in oligotrophic and eutrophic lakes, Annual Review of Ecology and Systematics, 9 (1978), 475–495. doi: 10.1146/annurev.es.09.110178.002355
|
| [25] | J. W. M. La Rivière, Microbial ecology of liquid waste treatment, in Advances in Microbial Ecology, vol. 1, Springer US, 1977,215–259. |
| [26] |
X. Li, J. Shen and R. Rakkiyappan, Persistent impulsive effects on stability of functional differential equations with finite or infinite delay, Applied Mathematics and Computation, 329 (2018), 14–22. doi: 10.1016/j.amc.2018.01.036
|
| [27] |
X. Li, X. Yang and T. Huang, Persistence of delayed cooperative models: Impulsive control method, Applied Mathematics and Computation, 342 (2019), 130–146. doi: 10.1016/j.amc.2018.09.003
|
| [28] | J. Monod, La technique de culture continue: Théorie et applications, Annales de l'Institute Pasteur, 79 (1950), 390–410. |
| [29] |
A. Novick and L. Szilard, Experiments with the chemostat on spontaneous mutations of bacteria, Proceedings of the National Academy of Sciences, 36 (1950), 708–719. doi: 10.1073/pnas.36.12.708
|
| [30] | S. Pilyugin and P. Waltman, The simple chemostat with wall growth, Siam Journal on Applied Mathematics - SIAMAM, 59. |
| [31] |
E. Rurangwa and M. C. J. Verdegem, Microorganisms in recirculating aquaculture systems and their management, Reviews in Aquaculture, 7 (2015), 117–130. doi: 10.1111/raq.12057
|
| [32] | H. L. Smith and P. Waltman, The theory of the chemostat: dynamics of microbial competition, Cambridge University Press, 1995. |
| [33] | V. Sree Hari Rao and P. Raja Sekhara Rao, Dynamic Models and Control of Biological Systems, Springer-Verlag, Heidelberg, 2009. |
| [34] |
G. Stephanopoulos, R. Aris and A. Fredrickson, A stochastic analysis of the growth of competing microbial populations in a continuous biochemical reactor, Mathematical Biosciences, 45 (1979), 99–135. doi: 10.1016/0025-5564(79)90098-1
|
| [35] |
F. Stewart and B. Levin, The population biology of bacterial plasmids: A priori conditions for the existence of conjugationally transmitted factors, Genetics, 87 (1977), 209–28. doi: 10.1093/genetics/87.2.209
|
| [36] |
P. A. Taylor and J. L. Williams, Theoretical studies on the coexistence of competing species under continuous flow conditions, Cadandian Journal of Microbiology, 21 (1975), 90–98. doi: 10.1139/m75-013
|
| [37] | H. R. Thieme, Convergence results and a poincare-bendixson trichotomy for asymptotically autonomous differential equations, Journal of Mathematical Biology, 30. |
| [38] |
J. López-de-la-Cruz, Random and stochastic disturbances on the input flow in chemostat models with wall growth, Stochastic Analysis and Applications, 37 (2019), 668–698. doi: 10.1080/07362994.2019.1605911
|
| [39] |
H. Veldcamp, Ecological studies with the chemostat, Advances in Microbial Ecology, 1 (1977), 59–95. doi: 10.1007/978-1-4615-8219-9_2
|
| [40] | P. Waltman, Competition Models in Population Biology, CBMS-NSF Regional Conference Series in Applied Mathematics, 1983, Society for Industrial and Applied Mathematics, Philadelphia. |
| [41] | P. Waltman, S. P. Hubbel and S. B. Hsu, Theoretical and experimental investigations of microbial competition in continuous culture, Modeling and Differential Equations in Biology (Conf., southern Illinois Univ. Carbonadle, III., 1978), 58 (1980), 107–152. |

























Figures(26)
Tomás Caraballo, Javier López-de-la-Cruz. Survey on chemostat models with bounded random input flow[J]. Mathematical Modelling and Control, 2021, 1(1): 52-78. doi: 10.3934/mmc.2021005







 DownLoad:
DownLoad: