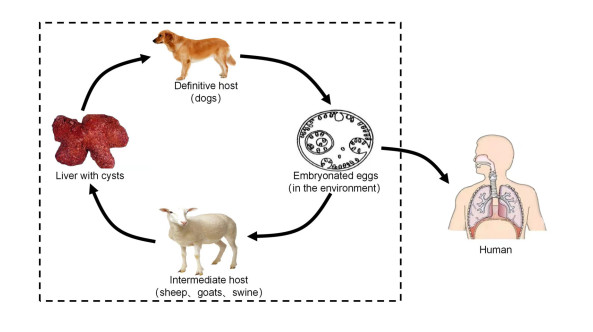
A transmission dynamics model with the logistic growth of cystic echinococcus in sheep was formulated and analyzed. The basic reproduction number was derived and the results showed that the global dynamical behaviors were determined by its value. The disease-free equilibrium is globally asymptotically stable when the value of the basic reproduction number is less than one; otherwise, there exists a unique endemic equilibrium and it is globally asymptotically stable. Sensitivity analysis and uncertainty analysis of the basic reproduction number were also performed to screen the important factors that influence the spread of cystic echinococcosis. Contour plots of the basic reproduction number versus these important factors are presented, too. The results showed that the higher the deworming rate of dogs, the lower the prevalence of echinococcosis in sheep and dogs. Similarly, the higher the slaughter rate of sheep, the lower the prevalence of echinococcosis in sheep and dogs. It also showed that the spread of echinococcosis has a close relationship with the maximum environmental capacity of sheep, and that they have a remarkable negative correlation. This reminds us that the risk of cystic echinococcosis may be underestimated if we ignore the increasing number of sheep in reality.
Citation: Yiwei He, Qianqian Cui, Zengyun Hu. Modeling and analysis of the transmission dynamics of cystic echinococcosis: Effects of increasing the number of sheep[J]. Mathematical Biosciences and Engineering, 2023, 20(8): 14596-14615. doi: 10.3934/mbe.2023653
A transmission dynamics model with the logistic growth of cystic echinococcus in sheep was formulated and analyzed. The basic reproduction number was derived and the results showed that the global dynamical behaviors were determined by its value. The disease-free equilibrium is globally asymptotically stable when the value of the basic reproduction number is less than one; otherwise, there exists a unique endemic equilibrium and it is globally asymptotically stable. Sensitivity analysis and uncertainty analysis of the basic reproduction number were also performed to screen the important factors that influence the spread of cystic echinococcosis. Contour plots of the basic reproduction number versus these important factors are presented, too. The results showed that the higher the deworming rate of dogs, the lower the prevalence of echinococcosis in sheep and dogs. Similarly, the higher the slaughter rate of sheep, the lower the prevalence of echinococcosis in sheep and dogs. It also showed that the spread of echinococcosis has a close relationship with the maximum environmental capacity of sheep, and that they have a remarkable negative correlation. This reminds us that the risk of cystic echinococcosis may be underestimated if we ignore the increasing number of sheep in reality.

| [1] |
C. M. Budke, P. Deplazes, P. R. Torgerson, Global socioeconomic impact of cystic echinococcosis, Emerging Infect. Dis., 12 (2006), 296–303. https://doi.org/10.3201/eid1202.050499 doi: 10.3201/eid1202.050499

|
| [2] | H. Wen, M. Q. Xu, Practical echinology (in Chinese), Science Press, 2007. |
| [3] |
D. J. Jenkins, WHO/OIE manual on echinococcosis in humans and animals: a public health problem of global concern, Int. J. Parasitol., 31 (2001), 1717–1718. https://doi.org/10.1016/S0020-7519(01)00318-6 doi: 10.1016/S0020-7519(01)00318-6
|
| [4] | D. P. McManus, W. Zhang, J. Li, P. B. Bartley, Echinococcosis, The lancet, 362 (2003), 1295–1304. https://doi.org/10.1016/S0140-6736(03)14573-4 |
| [5] |
Q. Yu, N. Xiao, S. Han, T. Tian, X. N. Zhou, Progress on the national echinococcosis control programme in China: analysis of humans and dogs population intervention during 2004–2014, Infect. Dis. Poverty, 9 (2020), 69–80. https://doi.org/10.1186/s40249-020-00747-7 doi: 10.1186/s40249-020-00747-7
|
| [6] |
P. R. Torgerson, The use of mathematical models to simulate control options for echinococcosis, Acta Trop., 85 (2003), 211–221. https://doi.org/10.1016/S0001-706X(02)00227-9 doi: 10.1016/S0001-706X(02)00227-9
|
| [7] |
P. R. Torgerson, D. D. Heath, Transmission dynamics and control options for echinococcus granulosus, Parasitology, 127 (2003), S143–S158. https://doi.org/10.1017/S0031182003003810 doi: 10.1017/S0031182003003810
|
| [8] |
P. R. Torgerson, B. S. Shaikenov, A. T. Rysmukhambetova, A. E. Ussenbayev, A. M. Abdybekova, K. K. Burtisurnov, Modelling the transmission dynamics of echinococcus granulosus in dogs in rural Kazakhstan, Parasitology, 126 (2003), 417–424. https://doi.org/10.1017/S0031182003002932 doi: 10.1017/S0031182003002932
|
| [9] |
M. G. Roberts, J. R. Lawson, M. A. Gemmell, Population dynamics in echinococcosis and cysticercosis: mathematical model of the life-cycles of Taenia hydatigena and T.ovis, Parasitology, 94 (1987), 181–197. https://doi.org/10.1017/S0031182000053555 doi: 10.1017/S0031182000053555
|
| [10] |
M. G. Roberts, J. R. Lawson, M. A. Gemmell, Population dynamics in echinococcosis and cysticercosis: mathematical model of the life-cycle of Echinococcus granulosus, Paraisitology, 92 (1986), 621–641. https://doi.org/10.1017/S0031182000065495 doi: 10.1017/S0031182000065495
|
| [11] |
M. A. Gemmell, J. R. Lawson, M. G. Roberts, Population dynamics in echinococcosis and cysticercosis: biological parameters of echinococcus granulosus in dogs and sheep, Parasitology, 92 (1986), 599–620. https://doi.org/10.1017/S0031182000065483 doi: 10.1017/S0031182000065483
|
| [12] |
M. A. Gemmell, J. R. Lawson, M. G. Roberts, B. R. Kerin, C. J. Mason, Population dynamics in echinococcosis and cysticercosis: comparison of the response of echinococcus granulosus, Taenia hydatigena and T.ovis to control, Parasitology, 93 (1986), 357–369. https://doi.org/10.1017/S0031182000051520 doi: 10.1017/S0031182000051520
|
| [13] |
M. A. Gemmell, J. R. Lawson, M. G. Roberts, Population dynamics in echinococcosis and cysticercosis: evaluation of the biological parameters of Taenia hydatigena and T.ovis and comparison with those of echinococcus granulosus, Parasitology, 94 (1987), 161–180. https://doi.org/10.1017/S0031182000053543 doi: 10.1017/S0031182000053543
|
| [14] |
K. Wang, X. Zhang, Z. Jin, H. Ma, Z. Teng, L. Wang, Modeling and analysis of the transmission of echinococcosis with application to Xinjiang Uygur Autonomous Region of China, J. Theor. Biol., 333 (2013), 78–90. https://doi.org/10.1016/j.jtbi.2013.04.020 doi: 10.1016/j.jtbi.2013.04.020
|
| [15] |
X. M. Rong, M. Fan, X. D. Sun, Y. M. Wang, H. P. Zhu, Impact of disposing stray dogs on risk assessment and control of echinococcosis in Inner Mongolia, Math. Biosci., 299 (2018), 85–96. https://doi.org/10.1016/j.mbs.2018.03.008 doi: 10.1016/j.mbs.2018.03.008
|
| [16] |
X. M. Rong, M. Fan, H. P. Zhu, Y. H. Zheng, Dynamic modeling and optimal control of cystic echinocococcosis, Infect. Dis. Poverty, 10 (2021), 1–13. https://doi.org/10.1186/s40249-021-00807-6 doi: 10.1186/s40249-021-00807-6
|
| [17] |
J. Zhao, R. Yang, A dynamical model of echinococcosis with optimal control and cost-effectiveness, Nonlinear Anal. Real World Appl., 62 (2021), 103388. https://doi.org/10.1016/j.nonrwa.2021.103388 doi: 10.1016/j.nonrwa.2021.103388
|
| [18] |
Q. Q. Cui, Modeling and analysis of the transmission dynamics of echinococcosis in Pengyang county, Ningxia (in Chinese), Math. Appl., 36 (2023), 277–289. https://doi.org/10.13642/j.cnki.42-1184/o1.2023.02.011 doi: 10.13642/j.cnki.42-1184/o1.2023.02.011
|
| [19] |
G. Zhu, S. Chen, B. Shi, H. Qiu, S. Xia, Dynamics of echinococcosis transmission among multiple species and a case study in Xinjiang, China, Chaos Solitons Fractals, 127 (2019), 103–109. https://doi.org/10.1016/j.chaos.2019.06.032 doi: 10.1016/j.chaos.2019.06.032
|
| [20] |
G. B. Birhan, J. M. W. Munganga, A. S. Hassan, Mathematical modeling of echinococcosis in humans, dogs, and sheep, J. Appl. Math., 2020 (2020), 1–18. https://doi.org/10.1155/2020/8482696 doi: 10.1155/2020/8482696
|
| [21] |
G. B. Bitew, J. M. W. Munganga, A. S. Hassan, Mathematical modelling of echinococcosis in human, dogs and sheep with intervention, J. Biol. Dyn., 16 (2022), 439–463. https://doi.org/10.1080/17513758.2022.2081368 doi: 10.1080/17513758.2022.2081368
|
| [22] |
S. Yang, W. Wu, T. Tian, J. Zhao, K. Chen, Q. Wang, et al., Prevalence of cystic echinococcosis in slaughtered sheep as an indicator to assess cntrol progress in Emin County, Xinjiang, China, Korean J. Parasitol., 53 (2015), 355–359. https://doi.org/10.3347/kjp.2015.53.3.355 doi: 10.3347/kjp.2015.53.3.355
|
| [23] |
G. L. Xiao, Q. Zhong, W. H. Xie, X. Wang, Epidemiological survey of sheep hydatidosis in Kashi Area of Xinjiang from 2014 to 2017 (in Chinese), China Anim. Health Insp., 36 (2019), 1–5. https://doi.org/10.3969/j.issn.1005-944X.2019.05.001 doi: 10.3969/j.issn.1005-944X.2019.05.001
|
| [24] |
Y. Gao, W. Wang, C. Lyu, X. Wei, Y. Chen, Q. Zhao, et al., Meta-analysis of the prevalence of echinococcus in sheep in China from 1983 to 2020, Front. Cell. Infect. Microbiol., 11 (2021), 668. https://doi.org/10.3389/fcimb.2021.711332 doi: 10.3389/fcimb.2021.711332
|
| [25] |
O. Diekmann, J. A. P. Heesterbeek, J. A. J. Metz, On the definition and the computation of the basic reproduction ratio $R_0$ in models for infectious diseases in heterogeneous populations, J. Math. Biol., 28 (1990), 365–382. https://doi.org/10.1007/BF00178324 doi: 10.1007/BF00178324
|
| [26] |
P. Dreessche, J. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180 (2002), 29–48. https://doi.org/10.1016/S0025-5564(02)00108-6 doi: 10.1016/S0025-5564(02)00108-6
|
| [27] | K. Dietz, Transmission and control of arbovirus diseases, Epidemiology, 104 (1975), 104–121. |
| [28] |
J. LaSalle, S. Lefschetz, R. C. Alverson, Stability by Liapunov's direct method with applications, Phys. Today, 15 (1962), 59. https://doi.org/10.1063/1.3057800 doi: 10.1063/1.3057800
|
| [29] | C. Castillo-Chavez, H. Thieme, Asymptotically autonomous epidemic models, in Mathematical Population Dynamics: Analysis of Heterogeneity, Springer, Berlin, (1995), 33–50. |
| [30] |
K. Mischaikow, H. Smith, H. R. Thieme, Asymptotically autonomous semiflows: chain recurrence and Lyapunov functions, Trans. Am. Math. Soc., 347 (1995), 1669–1685. https://doi.org/10.1090/S0002-9947-1995-1290727-7 doi: 10.1090/S0002-9947-1995-1290727-7
|
| [31] | Ningxia Hui Autonomous Region Statistics Bureau, Statistical yearbook, Available from: http://tj.nx.gov.cn/. |
| [32] | Q. R. Liu, X. L. Wu, R. Wen, Surveillance for echinococcosis in Pengyang county of Ningxia Hui Autonomous Region, 2011–2018, Dis. Surveillance, 35 (2020), 1125–1128. |
| [33] |
C. M. Budke, J. M. Qiu, Q. Wang, P. Torgerson, Economic effects of echinococcosis in a disease endemic region of the Tibetan Plateau, Am. J. Trop. Med. Hyg., 73 (2005), 2–10. https://doi.org/10.4269/ajtmh.2005.73.2 doi: 10.4269/ajtmh.2005.73.2
|
| [34] | Z. Y. Hu, D. L. Chen, X. Chen, Q. Zhou, Y. Peng, J. Li, et al., CCHZ-DISO: A timely new assessment system for data quality or model performance from Da Dao Zhi Jian, Geophys. Res. Lett., 49 (2022). https://doi.org/10.1029/2022GL100681 |








Figures(9) / Tables(1)

Yiwei He, Qianqian Cui, Zengyun Hu. Modeling and analysis of the transmission dynamics of cystic echinococcosis: Effects of increasing the number of sheep[J]. Mathematical Biosciences and Engineering, 2023, 20(8): 14596-14615. doi: 10.3934/mbe.2023653







 DownLoad:
DownLoad: