Citation: Kai Wang, Zhidong Teng, Xueliang Zhang. Dynamical behaviors of an Echinococcosis epidemic model with distributed delays[J]. Mathematical Biosciences and Engineering, 2017, 14(5&6): 1425-1445. doi: 10.3934/mbe.2017074

| [1] | [ Baidu, How Long is the Life Expectancy of Dogs? 2011, Available from: http://wenku.baidu.com/view/e7ccf4fec8d376eeaeaa31f9.html. |
| [2] | [ Baidu, The Sixth Census Data Announced by Xinjiang 2011, Available from: http://wenku.baidu.com/view/83e6d26cb84ae45c3b358cf1.html. |
| [3] | [ S. A. Berger and J. S. Marr, Human Parasitic Diseases Sourcebook, 1$^{st}$ edition, Jones and Bartlett Publishers: Sudbury, Massachusetts, 2006. |
| [4] | [ P. A. Cabrera,G. Haran,U. benavidez,S. Valledor,G. Perera,S. Lloyd,M. A. Gemmell,M. Baraibar,A. Morana,J. Maissonave,M. Carballo, Transmission dynamics of Echinococcus granulosus, Taenia hydatigena and Taenia ovis in sheep in Uruguay, Int. J. Parasitol., 25 (1995): 807-813. |
| [5] | [ Y. Cao,J. Wen,Q. Zheng, Analysis of the epidemic status of Echinococcosis in Xinjiang in 2010, J. Ningxia Med. Univ., 33 (2011): 784-788. |
| [6] | [ J. Eckert, M. A. Gemmell, F. X. Meslin and Z. S. Pawlowski, WHO/OIE manual on Echinococcosis in humans and animals: A public health problem of global concern, Paris: World Health Organization/World Organization for Animal Health, 2001. |
| [7] | [ Y. Kuang, Delay Differential Equations with Applications in Population Dynamics, Academic Press, NewYork, 1993. |
| [8] | [ S. Lahmar,H. Debbek,L. H. Zhang,D. P. McManus,A. Souissi,S. Chelly,P. R. Torgerson, Tansmission dynamics of the Echinococcus granulosus sheep-dog strain(G1 genotype) in camels in Tunisia, Vet. Parasitol., 121 (2004): 151-156. |
| [9] | [ Malike,Nusilaiti,Zulihumaer,Lv,Wali,Xi,Abudureyimu,Qiu,Abuduaini,Hanati, Investigation of Echinococcus infection in domestic in Xinjiang, Chin. J. Anim.Infect. Dis., 19 (2011): 57-60(in Chinese). |
| [10] | [ National Bureau of Statistics of China, China Statistical Yearbook, 2011 Available from: http://www.stats.gov.cn/tjsj/ndsj/2011/indexch.htm. |
| [11] | [ Nusilaiti,Zulihumaer, The infection situation of Echinococcosis and countermeasures of some counties and cities in xinjiang, Xinjiang Livestock Industry, 6 (2011): 45-46. |
| [12] | [ H. Smith, Monotone Dynamical Systems: An Introduction to the Theory of Competitive and Cooperative Systems, American Mathematical Society, 1995. |
| [13] | [ K. Takumi,A. Vires,M. Chu,J. Mulder,P. Teunis,J. Giessen, Evidence for an increasing presence of Echinococcus multilocularis in foxes in The Netherlands, Inter. J. Parasitol., 38 (2008): 571-578. |
| [14] | [ The Government of Xinjiang Uygur Autonomous Region of China, The improvement of people's living standard, 2010, Available from: http://www.xinjiang.gov.cn/2011/11/15/64.html. |
| [15] | [ H. R. Thieme, Convergence results and a Poincare-Bendixson trichotomy for asymptotically automous differential equations, J. Math. Biol., 30 (1992): 755-763. |
| [16] | [ P. R. Torgerson,K. K. Burtisurnov,B. S. Shaikenov,A. T. Rysmukhambetova,A. M. Abdybekova,A. E. Ussenbayev, Modelling the transmission dynamics of Echinococcus granulosus in sheep and cattle in Kazakhstan, Vet. Parasitol., 114 (2003): 143-153. |
| [17] | [ S. Wang and S. Ye, Textbook of Medical Microbiology and Parasitology, 1$^{st}$ edition, Science Press, Beijing, 2006. |
| [18] | [ Z. Wang,X.-Q. Zhao, Global dynamics of a time-delayed dengue transmission model, Can. Appl. Math. Q., 20 (2012): 89-113. |
| [19] | [ K. Wang,X. Zhang,Z. Jin,H. Ma,Z. Teng,L. Wang, Modeling and analysis of the transmission of Echinococcosis with application to Xinjiang Uygur Autonomous Region of China, J. Theor. Biol., 333 (2013): 78-90. |
| [20] | [ W. Wu, The Chinese National Plan for the Control Echincoccosis, Urumq: ISUOG's World Congress of Hydatidology Final Programme & Abstracts Book, 2011(in Chinese). |
| [21] | [ L. Wu,B. Song,W. Du,J. Lou, Mathematical modelling and control of Echinococcus in Qinghai province, China, Math. Biosci. Eng., 10 (2013): 425-444. |
| [22] | [ Xinjiang CDC, Information Center of Xinjiang Autonomous Region, 2012, Available from: http://www.xjcdc.com/. |
| [23] | [ Xi,Song,Xue,Nusilaiti,Malike,Zulihumaer, Survey on Echinococcus granulosus infection in shepherd dogs in tianshan mountainous area of Hejing city, Xinjiang province, Chin. J. Anim. Infect. Dis., 18 (2010): 59-62. |
| [24] | [ Z. Xu,X.-Q. Zhao, A vector-bias malaria model with incubation period and diffusion, Discrete Contin. Dyn. Syst. Ser. B, 17 (2012): 2015-2034. |
| [25] | [ S. Yan,Y. Zhang, The control and prevention of livestock Echinococcosis in Xinjiang, Grass-Feeding Livestock, null (1994): 45-47. |
| [26] | [ X.-Q. Zhao,Z. Jing, Global asymptotic behavior in some cooperative systems of functional differential equations, Canad. Appl. Math. Q., 4 (1996): 421-444. |
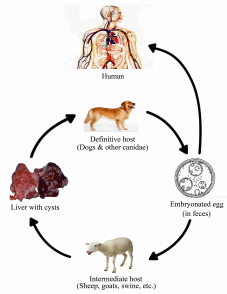
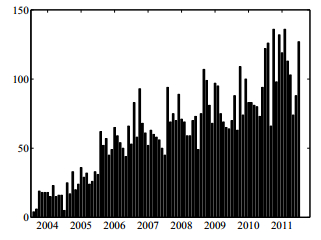
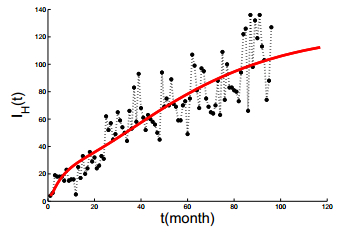
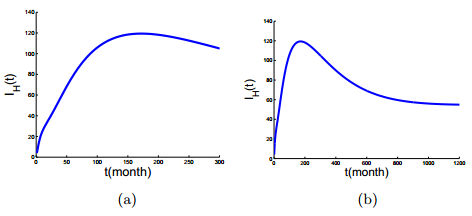
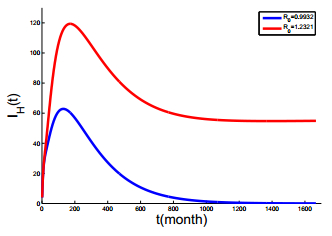
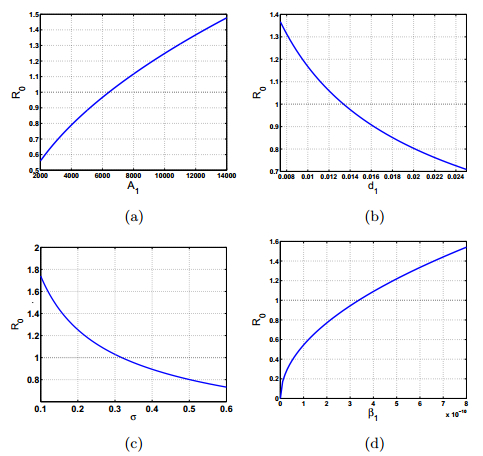
Figures(6) / Tables(3)

Kai Wang, Zhidong Teng, Xueliang Zhang. Dynamical behaviors of an Echinococcosis epidemic model with distributed delays[J]. Mathematical Biosciences and Engineering, 2017, 14(5&6): 1425-1445. doi: 10.3934/mbe.2017074







 DownLoad:
DownLoad: