Citation: Qiang Hou, Haiyan Qin. Global dynamics of a multi-stage brucellosis model with distributed delays and indirect transmission[J]. Mathematical Biosciences and Engineering, 2019, 16(4): 3111-3129. doi: 10.3934/mbe.2019154

| [1] | M. Madsen and E.C. Anderson, Serologic survey of Zimbabwean wildlife for brucellosis, J. Zoo Wildlife Med., 26 (1995), 240–245. |
| [2] | M.E. Meyer and M. Meagher, Brucellosis in freeranging bison (Bison bison) in Yellowstone, Grand Teton, and Wood Bu alo National Parks: A review, J. Wildlife Dis., 31 (1995), 579–598. |
| [3] | M. Mustafa and P. Nicoletti, Proceedings of the Workshop on Guidelines for a Regional Brucellosis Control Program for the Middle East 14-17 February, Amman, Jordan, FAO, WHO and OIE, 1993. |
| [4] | M. Darwish and A. Benkirane, Field investigations of brucellosis in cattle and small ruminants in Syria, 1990–1996, Rev. Sci. Tech. OIE, 20(3) (2001), 769–775. |
| [5] | J.R. Ebright, T. Altantsetseg and R. Oyungerel, Emerging infectious diseases in Mongolia, Emerg. Infect. Dis., 9(12) (2003), 1509–1515. |
| [6] | G. Pappas, P. Papadimitriou, N. Akritidis, et al., The new global map of human brucellosis, Lancet Infect. Dis., 6 (2006), 91–99. |
| [7] | M. Corbel, Brucellosis: An overview, Emerg. Infect. Dis., 3 (1997), 213–221. |
| [8] | J. McDermott, D. Grace and J. Zinsstag, Economics of brucellosis impact and control in lowincome countries, Rev. Sci. Tech., 32 (2013), 249–261. |
| [9] | M.J. Corbel, Brucellosis in humans and animals, WHO, FAO and OIE, 2006. |
| [10] | J. Parnas, W. Krüger and E. Tüppich, Die Brucellose des Menschen (Human Brucellosis), VEB VErlag Volk und Gesundheit, Berlin, 1966. |
| [11] | H. Krauss, A. Weber, B. Enders, et al., Zoonotic diseases, infection diseases transmitted from animals to human (Zoonosen, von Tier auf den Menschen übertragbare Infek-tionskrankheiten), Deutscher Ärzte-Verlag Köln, Cologne, 1996. |
| [12] | SCAHAW, Brucellosis in sheep and goats, Scientific Committee on Animal Health and Animal Welfare, 2001. |
| [13] | E.D. Ebel, M.S. Williams and S.M. Tomlinson, Estimating herd prevalence of bovine brucellosis in 46 U.S.A. states using slaughter surveillance, Prev. Vet. Med., 85 (2008), 295–316. |
| [14] | H.S. Lee, M. Her, M. Levine, et al. Time series analysis of human and bovine brucellosis in South Korea from 2005 to 2010, Prev. Vet. Med., 110 (2013), 190–197. |
| [15] | R.D. Jones, L. Kelly, T. England, et al., A quantitative risk assessment for the importation of brucellosis-infected breeding cattle into Great Britain from selected European countries, Prev. Vet. Med., 63 (2004), 51–61. |
| [16] | A.M. Al-Majali and M. Shorman, Childhood brucellosis in Jordan: Prevalence and analysis of risk factors, Int. J. Infect. Dis., 13 (2009), 196–200. |
| [17] | P. Jia and A. Joyner, Human brucellosis occurrences in inner mongolia, China: A spatio-temporal distribution and ecological niche modeling approach, BMC Infect. Dis., 13(36) (2015). |
| [18] | J. González-Guzmán and R. Naulin, Analysis of a model of bovine brucellosis using singular perturbations, J. Math. Biol., 33 (1994), 211–234. |
| [19] | J.Zinsstag, F. Roth, D. Orkhon, et al., A model of animal-human brucellosis transmission in Mongolia, Prev. Vet. Med., 69 (2005), 77–95. |
| [20] | D. Andrew and M. Mary, The Population Dynamics of Brucellosis in the Yellowstone National Park, Ecology, 77(4) (1996), 1026–1036. |
| [21] | F. Xie and R.D. Horan, Disease and Behavioral Dynamics for Brucellosis Control in Elk and Cattle in the Greater Yellowstone Area, J. Agr. Resour. Econ., 34 (2009), 11–33. |
| [22] | D. Shabb, N. Chitnis, Z. Baljinnyam, et al., A mathematical model of the dynamics of Mongolian livestock populations, Livest. Sci., 157 (2013), 280–288. |
| [23] | B. AÏnseba, C. Benosman and P. Magal, A model for ovine brucellosis incorporating direct and indirect transmission, J. Biol. Dyn., 4(1) (2010), 2–11. |
| [24] | M.T. Li, G.Q. Sun, J. Zhang, et al., Transmission dynamics and control for a brucellosis model In Hinggan League of Inner Mongolia, China, Math. Biosci. Eng., 11 (2014), 1115–1137. |
| [25] | Q. Hou, X.D Sun, J. Zhang, et al., Modeling the Transmission Dynamics of Sheep Brucellosis in Inner Mongolia Autonomous Region, China, Math. Biosci., 242 (2013), 51–58. |
| [26] | Q. Hou, X.D. Sun, Y.M. Wang, et al., Global properties of a general dynamic model for animal diseases: A case study of brucellosis and tuberculosis transmission, J. Math. Anal. Appl., 414(1) (2014), 424–433. |
| [27] | P.O. Lolika, C. Modnak and S. Mushayabasa, On the dynamics of brucellosis infection in bison population with vertical transmission and culling, Math. Biosci., 305 (2018), 42–54. |
| [28] | W.M. Liu, S.A. Levin and Y. Iwasa, Influence of nonlinear incidence rates upon the behavior of SIRS epidemiological models, J. Math. Biol., 23 (1986), 187–204. |
| [29] | H. McCallum, N. Barlow and J. Hone, How should pathogen transmission be modelled ?, Trends Ecol. Evol., 16(6) (2001), 295–300. |
| [30] | R. Breban, J.M Drake, D.E. Stallknecht, et al., The role of environmental transmission in recurrent avian influenza epidemics, PLoS Comput. Biol., 5(4) (2009), e1000346. |
| [31] | Z. Mukandavire, S. Liao, J. Wang, et al., Estimating the reproductive numbers for the 2008–2009 cholera outbreaks in Zimbabwe, Proc. Natl. Acad. Sci. USA, 108 (2011), 8767–8772. |
| [32] | H.B. Guo, M.Y. Li and Z.S. Shuai, Global dynamics of a general class of multistage models for infectious diseases, SIAM J. Appl. Math., 72(1) (2012), 261–279. 33. J.P. LaSalle, Stability of Dynamical Systems, SIAM, Philadelphia, 1976. |
| [33] | 34. J.K. Hale and P. Waltman, Persistence in infinite-dimensional systems, SIAM J. Math. Anal., 20 (1989), 388–395. |
| [34] | 35. Y. Yang, L. Zou and S. Ruan, Global dynamics of a delayed within-host viral infection model with both virus-to-cell and cell-to-cell transmissions, Math. Biosci., 270 (2015), 183–191. |
| [35] | 36. O.P. Lolika and S. Mushayabasa, Dynamics and stability analysis of a brucellosis model with two discrete delays, Discrete Dyn. Nat. Soc., 2 (2018), 1–20. |
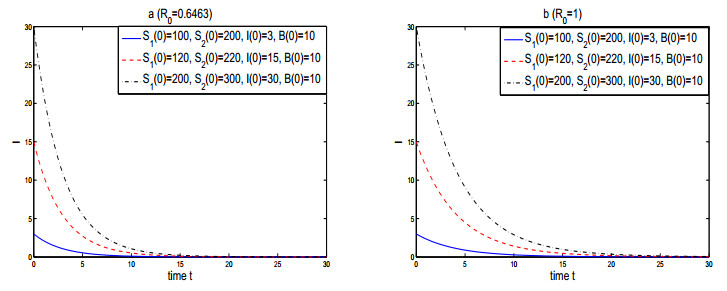
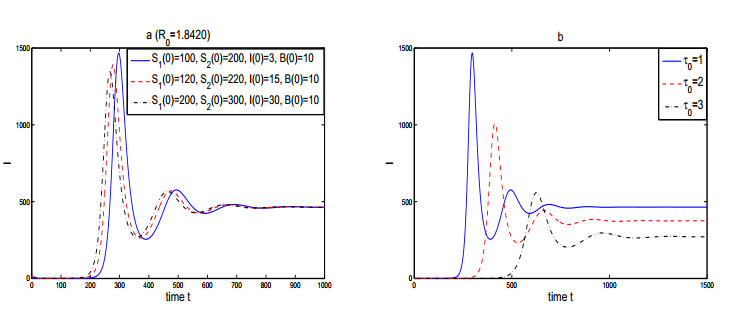
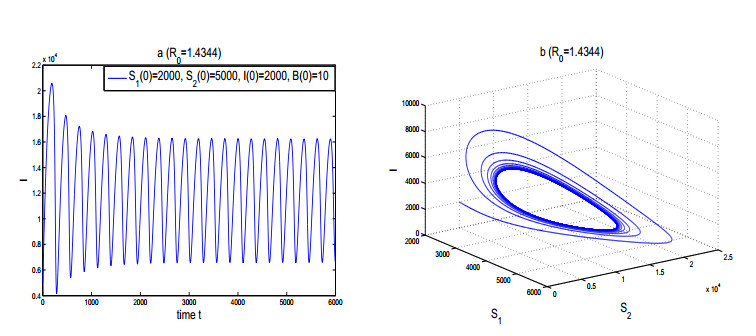
Figures(3)

Qiang Hou, Haiyan Qin. Global dynamics of a multi-stage brucellosis model with distributed delays and indirect transmission[J]. Mathematical Biosciences and Engineering, 2019, 16(4): 3111-3129. doi: 10.3934/mbe.2019154







 DownLoad:
DownLoad: