Citation: Mingtao Li, Xin Pei, Juan Zhang, Li Li. Asymptotic analysis of endemic equilibrium to a brucellosis model[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 5836-5850. doi: 10.3934/mbe.2019291

| [1] | M. J. Corbel, Brucellosis: an overview, Emerg. Infect. Dis., 3(1997), 213–221. |
| [2] | G. Pappas, N. Akritidis, M. Bosilkovski, et al., Brucellosis, N. Engl. J. Med., 352(2005), 2325–2536. |
| [3] | M. T. Li, G. Q. Sun, J. Zhang, et al., Transmission dynamics and control for a Brucellosis Model in Hinggan League of Inner Mongolia, China, Math. Biosci. Eng., 11(2014), 1115–1137. |
| [4] | M. L. Boschiroli, V. Foulongne and D. O'Callaghan, Brucellosis: a worldwide zoonosis, Curr. Opin. Microbiol., 4(2001), 58–64. |
| [5] | G. Pappas, P. Papadimitriou, N. Akritidis, et al., The new global map of human brucellosis, Lancet Infect. Dis., 6(2006), 91–99. |
| [6] | M. P. Franco, M. Mulder, R. H. Gilman, et al., Human brucellosis, Lancet Infect. Dis., 7(2007), 775–786. |
| [7] | M. N. Seleem, S. M. Boyle and N. Sriranganathan, Brucellosis: A re-emerging zoonosis, Vet. Microbiol., 140 (2010), 392–398. |
| [8] | H. Heesterbeek, R. M. Anderson, V. Andreasen, et al., Modeling infectious disease dynamics in the complex landscape of global health, Science, 6227 (2015), aaa4339. |
| [9] | M. T. Li, G. Q. Sun, J. Zhang, et al., Transmission dynamics of a multi-group brucellosis model with mixed cross infection in public farm, Appl. Math. Comput., 237(2014), 582–594. |
| [10] | G. G. Jorge and N. Raul, Analysis of a model of bovine brucellosis using singular perturbations, J. Math. Biol., 33(1994), 211–223. |
| [11] | J. Zinsstag, F. Roth, D. Orkhon, et al., A model of animalChuman brucellosis transmission in Mongolia, Prev. Vet. Med., 69(2005), 77–95. |
| [12] | B. Alnseba, B. Chahrazed and M. Pierre, A model for ovine brucellosis incorporating direct and indirect transmission, J. Biol. Dyn., 4(2010), 2–11. |
| [13] | Q. Hou, X. D. Sun, J. Zhang, et al., Modeling the transmission dynamics of brucellosis in Inner Mongolia Autonomous Region, China, Math. Biosci., 242(2013), 51–58. |
| [14] | Q. Hou, X. D. Sun, Y. M. Wang, et al., Global properties of a general dynamic model for animal diseases: A case study of brucellosis and tuberculosis transmission, J. Math. Anal. Appl., 414 (2014), 424–433. |
| [15] | W. Beauvais, I. Musallam and J. Guitian, Vaccination control programs for multiple livestock host species: An age-stratified, seasonal transmission model for brucellosis control in endemic settings, Paras. Vector, 9(2016), 55. |
| [16] | P. Lou, L. Wang, X. Zhang, et al., Modelling Seasonal Brucellosis Epidemics in Bayingolin Mongol Autonomous Prefecture of Xinjiang, China, 2010-2014, BioMed Res. Int., 2016(2016), 5103718. |
| [17] | D. O. Montiel, M. Bruce, K. Frankena, et al., Financial analysis of brucellosis control for small-scale goat farming in the Bajio region, Mexico, Prev. Vet. Med., 118(2015), 247–259. |
| [18] | L. Yang, Z. W. Bi, Z. Q. Kou, et al., Time-series analysis on human brucellosis during 2004-2013 in Shandong province, China, Zoonoses Public Health, 62(2015), 228–235. |
| [19] | M. T. Li, G. Q. Sun, W. Y. Zhang, et al., Model-based evaluation of strategies to control brucellosis in China, Int. J. Env. Res. Pub. He., 14(2017), 295. |
| [20] | M. T. Li, Z. Jin, G. Q. Sun, et al., Modeling direct and indirect disease transmission using multi-group model, J. Math. Anal. Appl., 446(2017), 1292–1309. |
| [21] | O. Diekmann, J. A. P. Heesterbeek and J. A. J. Metz, On the definition and the computation of the basic reproduction ratio R 0 in models for infectious diseases in heterogeneous populations, J. Math. Biol., 28(1990), 365–382. |
| [22] | P. van den Driessche and J. Watmough, Reproduction numbers and sub-threshold endemic equi-libria for compartmental models of disease transmission, Math. Biosci., 180(2002), 29–48. |
| [23] | O. Diekmann, J. A. P. Heesterbeek and M. G. Roberts, The construction of next-generation matrices for compartmental epidemic models, J. R. Soc. Interface, 7(2010), 873–885. |
| [24] | M. Y. Li and Z. Shuai, Global-stability problem for coupled systems of differential equations on networks, J. Diff. Equat., 248(2010), 1–20. |
| [25] | J. P. Lasalle, The stability of dynamical dystems, in: Regional Conference Series in Applied Mathematics, SIAM, Philadelphia, 1976. |
| [26] | M. Y. Li and Z. Shuai, Global stability of the endemic equilibrium of multigroup SIR epidemic models, Canad. Appl. Math. Quart., 14(2006), 259–284. |
| [27] | H. Guo, M. Y. Li and Z. Shuai, A graph-theoretic approach to the method of global lyapunov functions, Proc. Amer. Math. Soc., 136(2008), 2793–2802. |
| [28] | S. Marino, I. B. Hogue, C. J. Ray, et al., A methodology for performing global uncertainty and sensitivity analysis in systems biology, J. Theoret. Biol., 254(2008), 178–196. |
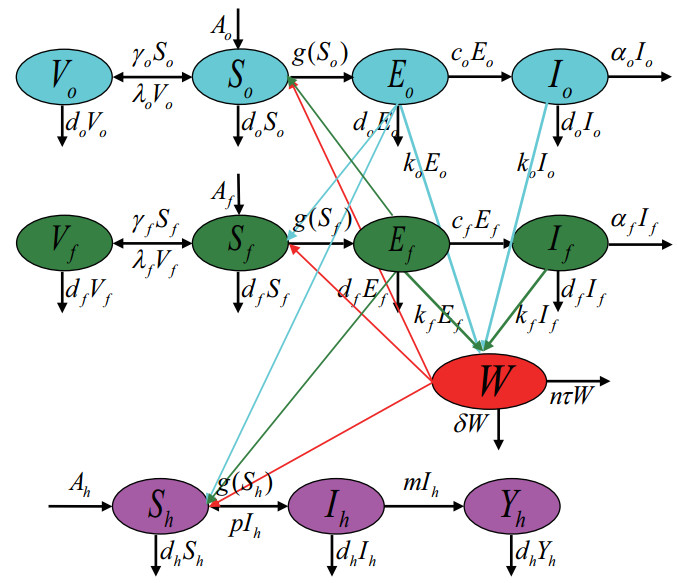
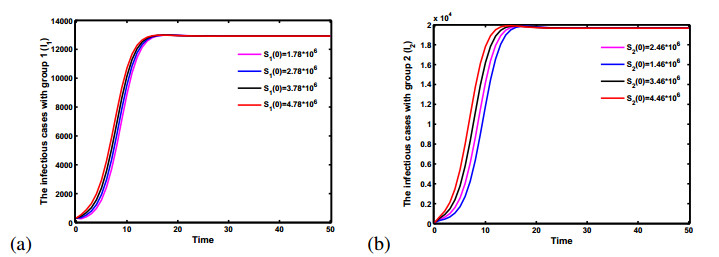
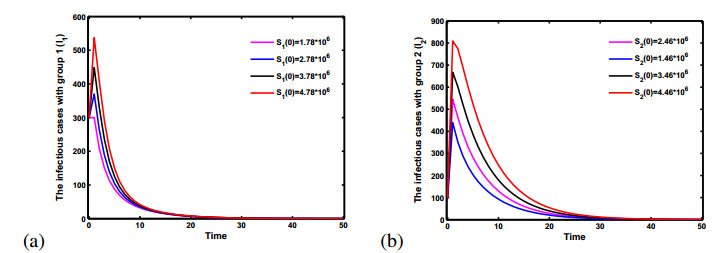
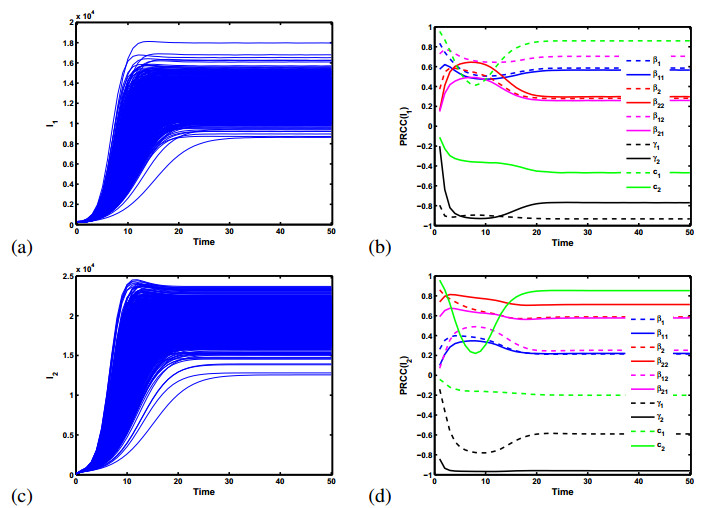
Figures(5)

Mingtao Li, Xin Pei, Juan Zhang, Li Li. Asymptotic analysis of endemic equilibrium to a brucellosis model[J]. Mathematical Biosciences and Engineering, 2019, 16(5): 5836-5850. doi: 10.3934/mbe.2019291







 DownLoad:
DownLoad: