Citation: Liqiong Pu, Zhigui Lin. A diffusive SIS epidemic model in a heterogeneous and periodically evolvingenvironment[J]. Mathematical Biosciences and Engineering, 2019, 16(4): 3094-3110. doi: 10.3934/mbe.2019153

| [1] | L. J. S. Allen, B. M. Bolker, Y. Lou, et al., Asymptotic profiles of the steady states for an SIS epidemic reaction-diffusion model, Discrete Contin. Dyn. Syst. Ser. A, 21 (2008), 1–20. |
| [2] | E. J. Crampin, Reaction Diffusion Patterns on Growing Domains, PhD thesis, University of Oxford, 2000. |
| [3] | E. J. Crampin, E. A. Gaffney and P. K. Maini, Reaction and diffusion on growing domains: scenarios for robust pattern formation, Bull. Math. Biol., 61 (1999), 1093–1120. 222 |
| [4] | E. J. Crampin, E. A. Gaffney and P. K. Maini Mode-doubling and tripling in reaction-diffusion patterns on growing domains: a piecewise linear model, J. Math. Biol., 44 (2002), 107–128. |
| [5] | E. J. Crampin, W. W. Hackborn and P. K. Maini, Pattern formation in reaction diffusion models with nonuniform domain growth, Bull. Math. Biol., 64 (2002), 747–769. |
| [6] | E. J. Crampin and P. K. Maini, Modelling biological pattern formation: the role of domain growth, Comm. Theor. Biol., 6 (2001), 229–249. |
| [7] | P. van den Driessche and J. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180 (2002), 29–48. |
| [8] | Y. Du and Z. Lin, Spreading-vanishing dichotomy in the diffusive logistic model with a free boundary, SIAM J. Math. Anal., 42 (2010), 377-405. |
| [9] | J. Ge, K. I. Kim, Z. G. Lin, et al., A SIS reaction-diffusion-advection model in a low-risk and high-risk domain, J. Differential Equations, 259 (2015), 5486-5509. |
| [10] | H. Gu, Z. Lin and B. Lou, Different asymptotic spreading speeds induced by advection in a diffusion problem with free boundaries, Proc. Amer. Math. Soc., 143 (2015), 1109–1117. |
| [11] | S. B. Hsu and Y. Lou, Single phytoplankton species growth with light and advection in a water column, SIAM J. Appl. Math., 70 (2010), 2942–2974. |
| [12] | W. Huang, M. Han and K. Liu, Dynamics of an SIS reaction-diffusion epidemic model for disease transmission, Math. Biosci. Eng., 7 (2010), 51–66. |
| [13] | D. H. Jiang and Z. C. Wang, The diffusive logistic equation on periodically evolving domains, J. Math. Anal. Appl. 458 (2018), 93–111. |
| [14] | C. X. Lei, Z. G. Lin and Q. Y. Zhang, The spreading front of invasive species in favorable habitat or unfavorable habitat, J. Differential Equations, 257 (2014), 145–166. |
| [15] | C. X. Lei, H. Nie, W. Dong, et al., Spreading of two competing species governed by a free boundary model in a shifting environment, J. Math. Anal. Appl., 462 (2018), 1254–1282. |
| [16] | H. C. Li, R. Peng and F. B.Wang, Varying total population enhances disease persistence: Qualitative analysis on a diffusive SIS epidemic model, J. Differential Equations, 262 (2017), 885–913. |
| [17] | Z. G. Lin and H. P. Zhu, Spatial spreading model and dynamics of West Nile virus in birds and mosquitoes with free boundary, J. Math. Biol., 75 (2017), 1381–1409. |
| [18] | C. V. Pao, Stability and attractivity of periodic solutions of parabolic systems with time delays, J. Math. Anal. Appl., 304 (2005), 423–450. |
| [19] | T. J. Poole and M. S. Steinberg, Amphibian pronephric duct morphogenesis: segregation, cell rearrangement and directed migration of the Ambystoma duct rudiment, Development, 63 (1981), 1–16. |
| [20] | S. Sun, L. Pu and Z. Lin, Dynamics of the logistic harvesting model with infinite delay on periodically evolving domains, Commun. Math. Biol. Neurosci., 2018 (2018), 19 pages. |
| [21] | Q. Tang and Z. Lin, The asymptotic analysis of an insect dispersal model on a growing domain, J. Math. Anal. Appl., 378 (2011), 649–656. |
| [22] | Q. Tang, L. Zhang and Z. Lin, Asymptotic profile of species migrating on a growing habitat, Acta Appl. Math., 116 (2011), 227. |
| [23] | C. Tian and S. Ruan, A free boundary problem for Aedes aegypti mosquito invasion, Appl. Math. Model., 46 (2017), 203–217. |
| [24] | M. Wang, The diffusive logistic equation with a free boundary and sign-changing coefficient, J. Differential Equations, 258 (2015), 1252–1266. |
| [25] | M. Wang and Y. Zhang, Dynamics for a diffusive prey-predator model with different free boundaries, J. Differential Equations, 264 (2018), 3527–3558. |
| [26] | W. D.Wang and X. Q. Zhao, Basic reproduction numbers for reaction-diffusion epidemic models, SIAM J. Appl. Dyn. Syst., 11 (2012), 1652-1673. |
| [27] | N. Waterstraat, On bifurcation for semilinear elliptic Dirichlet problems on shrinking domains, Springer Proc. Math. Stat., 415 (2014), 240–246. |
| [28] | X. Q. Zhao, Dynamical System in Population Biology, Second Edition, CMS Books in Mathematics /Ouvrage de Mathmatiques de la SMC, Springer, Cham, (2017). |
| [29] | Y. Zhao and M. Wang, A reaction-diffusion-advection equation with mixed and free boundary conditions, J. Dyn. Differ. Equ., 30 (2018), 743–777. |
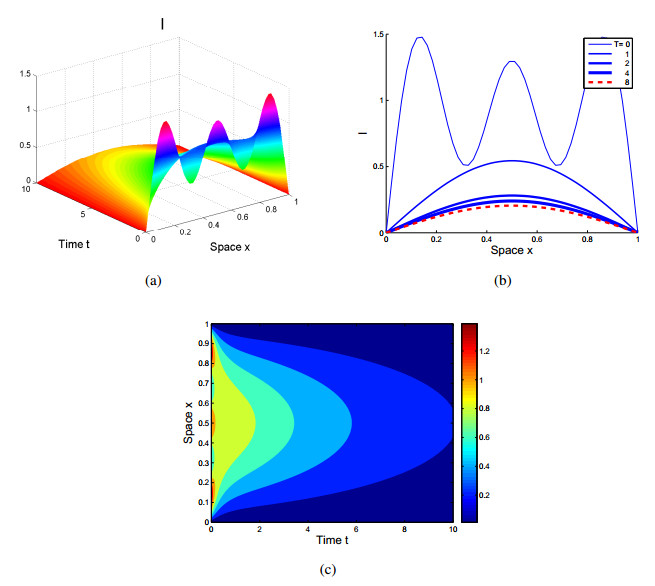
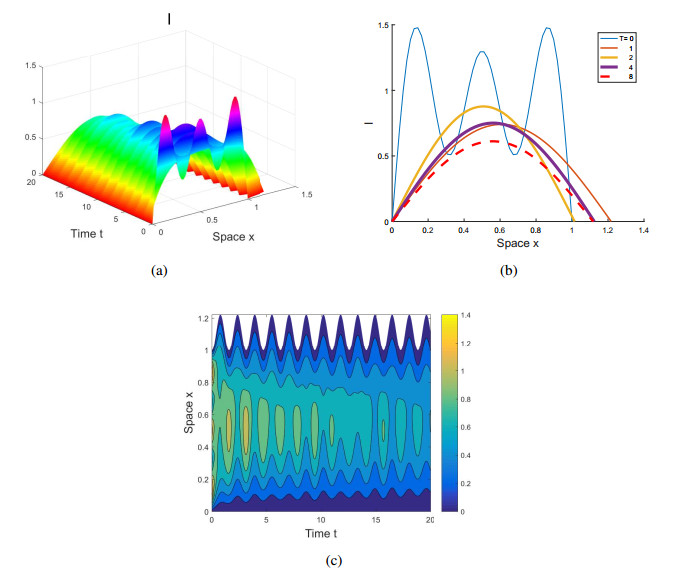
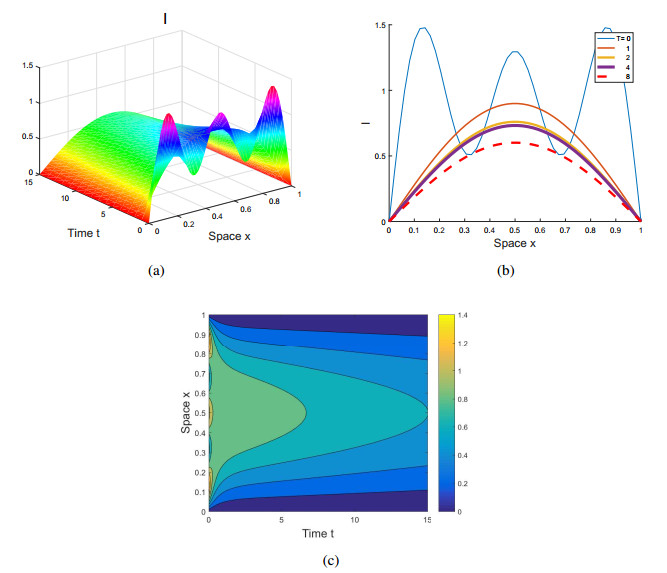
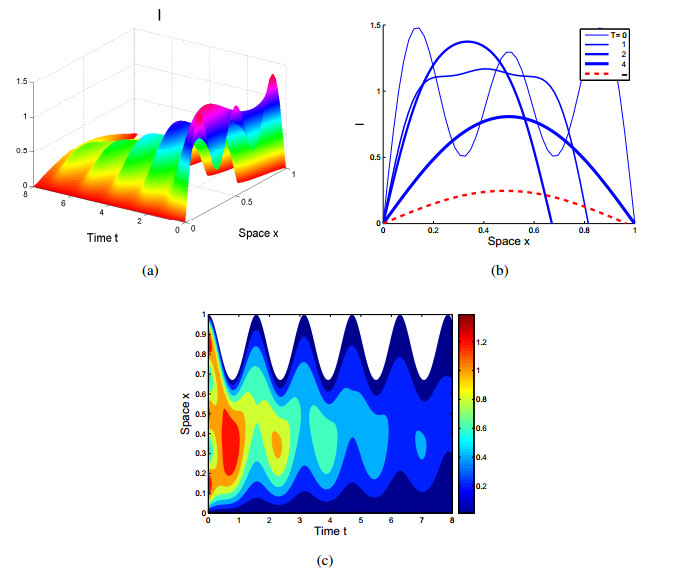
Figures(4)

Liqiong Pu, Zhigui Lin. A diffusive SIS epidemic model in a heterogeneous and periodically evolvingenvironment[J]. Mathematical Biosciences and Engineering, 2019, 16(4): 3094-3110. doi: 10.3934/mbe.2019153







 DownLoad:
DownLoad: