Citation: Xia Wang, Yuming Chen. An age-structured vector-borne disease model with horizontal transmission in the host[J]. Mathematical Biosciences and Engineering, 2018, 15(5): 1099-1116. doi: 10.3934/mbe.2018049

| [1] | [ C. J. Browne,S. S. Pilyugin, Global analysis of age-structured within-host virus model, Discrete Contin. Dyn. Syst. Ser. B, 18 (2013): 1999-2017. |
| [2] | [ Y. Chen,S. Zou,J. Yang, Global analysis of an SIR epidemic model with infection age and saturated incidence, Nonlinear Anal. Real World Appl., 30 (2016): 16-31. |
| [3] | [ K. Dietz, L. Molineaux and A. Thomas, A malaria model tested in the African savannah, Bull. World Health Organ., 50(1974), 347-357. |
| [4] | [ X. Feng,S. Ruan,Z. Teng,K. Wang, Stability and backward bifurcation in a malaria transmission model with applications to the control of malaria in China, Math. Biosci., 266 (2015): 52-64. |
| [5] | [ Z. Feng,J. X. Velasco-HerNández, Competitive exclusion in a vector-host model for the dengue fever, J. Math. Biol., 35 (1997): 523-544. |
| [6] | [ F. Forouzannia,A. B. Gumel, Mathematical analysis of an age-structured model for malaria transmission dynamics, Math. Biosci., 247 (2014): 80-94. |
| [7] | [ J. K. Hale, Asymptotic Behavior of Dissipative Systems, Am. Math. Soc., Providence, RI, 1988. |
| [8] | [ H. W. Hethcote, The mathematics of infectious diseases, SIAM Rev., 42 (2000): 599-653. |
| [9] | [ M. Iannelli, Mathematical Theory of Age-Structured Population Dynamics, Giardini Editori E Stampatori, Pisa, 1995. |
| [10] | [ H. Inaba,H. Sekine, A mathematical model for Chagas disease with infection-age-dependent infectivity, Math. Biosci., 190 (2004): 39-69. |
| [11] | [ Y. Kuang, Delay Differential Equations: With Applications in Population Dynamics, Academic Press, Boston, MA, 1993. |
| [12] | [ A. A. Lashari,G. Zaman, Global dynamics of vector-borne diseases with horizontal transmission in host population, Comput. Math. Appl., 61 (2011): 745-754. |
| [13] | [ Y. Lou,X.-Q. Zhao, A climate-based malaria transmission model with structured vector population, SIAM J. Appl. Math., 70 (2010): 2023-2044. |
| [14] | [ G. Macdonald, The analysis of equilibrium in malaria, Trop. Dis. Bull., 49 (1952): 813-829. |
| [15] | [ P. Magal,C. C. McCluskey,G. F. Webb, Lyapunov functional and global asymptotic stability for an infection-age model, Appl. Anal., 89 (2010): 1109-1140. |
| [16] | [ A. V. Melnik,A. Korobeinikov, Lyapunov functions and global stability for SIR and SEIR models with age-dependent susceptibility, Math. Biosci. Eng., 10 (2013): 369-378. |
| [17] | [ V. N. Novosltsev, A. I. Michalski, J. A. Novoseltsevam A. I. Tashin, J. R. Carey and A. M. Ellis, An age-structured extension to the vectorial capacity model, PloS ONE, 7 (2012), e39479. |
| [18] | [ Z. Qiu, Dynamical behavior of a vector-host epidemic model with demographic structure, Comput. Math. Appl., 56 (2008): 3118-3129. |
| [19] | [ R. Ross, The Prevention of Malaria, J. Murray, London, 1910. |
| [20] | [ R. Ross, Some quantitative studies in epidemiology, Nature, 87 (1911): 466-467. |
| [21] | [ S. Ruan,D. Xiao,J. C. Beier, On the delayed Ross-Macdonald model for malaria transmission, Bull. Math. Biol., 70 (2008): 1098-1114. |
| [22] | [ H. R. Thieme, Uniform persistence and permanence for non-autonomous semiflows in population biology, Math. Biosci., 166 (2000): 173-201. |
| [23] | [ J. Tumwiine,J. Y. T. Mugisha,L. S. Luboobi, A mathematical model for the dynamics of malaria in a human host and mosquito vector with temporary immunity, Appl. Math. Comput., 189 (2007): 1953-1965. |
| [24] | [ C. Vargas-de-León, Global analysis of a delayed vector-bias model for malaria transmission with incubation period in mosquitoes, Math. Biosci. Eng., 9 (2012): 165-174. |
| [25] | [ C. Vargas-de-León,L. Esteva,A. Korobeinikov, Age-dependency in host-vector models: The global analysis, Appl. Math. Comput., 243 (2014): 969-981. |
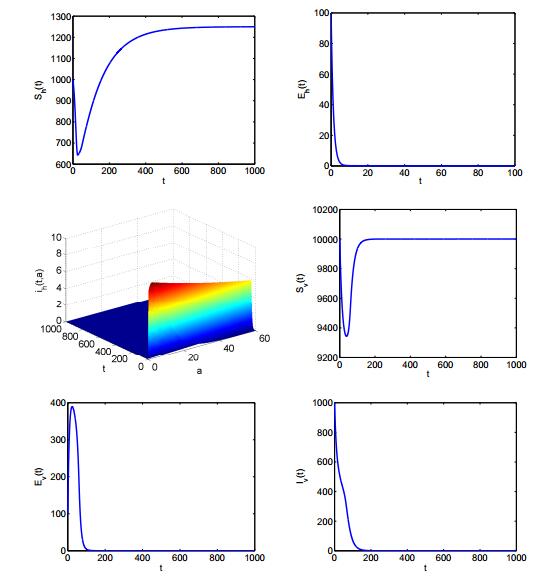
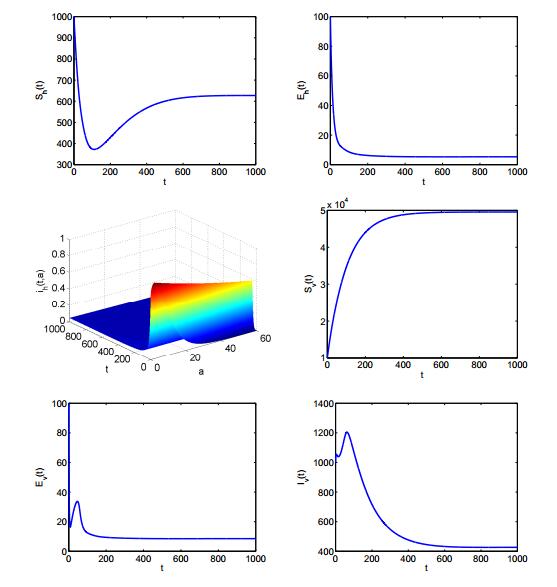
Figures(2) / Tables(1)

Xia Wang, Yuming Chen. An age-structured vector-borne disease model with horizontal transmission in the host[J]. Mathematical Biosciences and Engineering, 2018, 15(5): 1099-1116. doi: 10.3934/mbe.2018049







 DownLoad:
DownLoad: