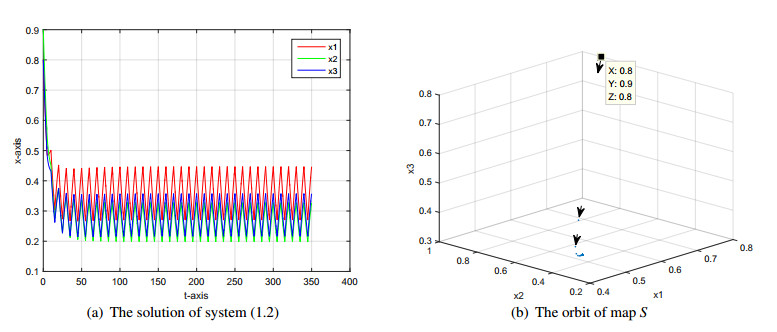
In this paper, we investigate a tridiagonal three-species competition model with seasonal succession. The Floquet multipliers of all nonnegative periodic solutions of such a time-periodic system are estimated via the stability analysis of equilibria. Together with the Brouwer degree theory, sufficient conditions for existence and uniqueness of the positive periodic solution are given. We further obtain the global dynamics of coexistence and extinction for three competing species in this periodically forced environment. Finally, some numerical examples are presented to illustrate the effectiveness of our theoretical results.
Citation: Xizhuang Xie, Meixiang Chen. Global stability of a tridiagonal competition model with seasonal succession[J]. Mathematical Biosciences and Engineering, 2023, 20(4): 6062-6083. doi: 10.3934/mbe.2023262
In this paper, we investigate a tridiagonal three-species competition model with seasonal succession. The Floquet multipliers of all nonnegative periodic solutions of such a time-periodic system are estimated via the stability analysis of equilibria. Together with the Brouwer degree theory, sufficient conditions for existence and uniqueness of the positive periodic solution are given. We further obtain the global dynamics of coexistence and extinction for three competing species in this periodically forced environment. Finally, some numerical examples are presented to illustrate the effectiveness of our theoretical results.

| [1] | H. L. Smith, Monotone Dynamical Systems. An Introduction to the Theory of Competitive and Cooperative Systems, American Mathematical Society, 1995. https://doi.org/10.1090/surv/041 |
| [2] | J. D. Murray, Mathematical Biology I: An Introduction. 3$^{rd}$ edition, Springer-Verlag, Berlin, 2002. |
| [3] | M. W. Hirsch, H. L. Smith, Monotone dynamical systems, in Handbook of Differential Equations: Ordinary Differential Equations, Elsevier, Amsterdam, 2005. https://doi.org/10.1016/S1874-5725(05)80006-9 |
| [4] | X. Q. Zhao, Dynamical systems in population biology. 2$^{nd}$ edition, Springer, New York, 2017. https://doi.org/10.1007/978-3-319-56433-3 |
| [5] |
J. Smillie, Competitive and cooperative tridiagonal systems of differential equations, SIAM J. Math. Anal., 15 (1984), 530–534. https://doi.org/10.1137/0515040 doi: 10.1137/0515040

|
| [6] |
H. L. Smith, Periodic tridiagonal competitive and cooperative systems of differential equations, SIAM J. Math. Anal., 22(1991), 1102–1109. https://doi.org/10.1137/0522071 doi: 10.1137/0522071
|
| [7] |
M. Gyllenberg, Y. Wang, Periodic tridiagonal systems modeling competitive-cooperative ecological interactions, Discrete Contin. Dyn. Syst. B, 5 (2005), 289–298. https://doi.org/10.3934/dcdsb.2005.5.289 doi: 10.3934/dcdsb.2005.5.289
|
| [8] |
C. Fang, M. Gyllenberg, Y. Wang, Floquet bundles for tridiagonal competitive-cooperative systems and the dynamics of time-recurrent systems, SIAM J. Math. Anal., 45 (2013), 2477–2498. https://doi.org/10.1137/120878021 doi: 10.1137/120878021
|
| [9] |
C. A. Klausmeier, Successional state dynamics: A novel approach to modeling nonequilibrium foodweb dynamics, J. Theor. Biol., 262 (2010), 584–595. https://doi.org/10.1016/j.jtbi.2009.10.018 doi: 10.1016/j.jtbi.2009.10.018
|
| [10] |
C. F. Steiner, A. S. Schwaderer, V. Huber, C. A. Klausmeier, E. Litchman, Periodically forced food chain dynamics: model predictions and experimental validation, Ecology, 90 (2009), 3099–3107. https://doi.org/10.1890/08-2377.1 doi: 10.1890/08-2377.1
|
| [11] |
S. B. Hsu, X. Q. Zhao, A Lotka-Volterra competition model with seasonal succession, J. Math. Biol., 64 (2012), 109–130. https://doi.org/10.1007/s00285-011-0408-6 doi: 10.1007/s00285-011-0408-6
|
| [12] |
L. Niu, Y. Wang, X. Xie, Carrying simplex in the Lotka-Volterra competition model with seasonal succession with applications, Discrete Contin. Dyn. Syst. B, 26 (2021), 2161–2172. https://doi.org/10.3934/dcdsb.2021014 doi: 10.3934/dcdsb.2021014
|
| [13] |
X. Xie, L. Niu, Global stability in a three-species Lotka-Volterra cooperation model with seasonal succession, Math. Meth. Appl. Sci., 44 (2021), 14807–14822. https://doi.org/10.1002/mma.7744 doi: 10.1002/mma.7744
|
| [14] |
Y. Zhang, X. Q. Zhao, Bistable travelling waves for reaction and diffusion model with seasonal succession, Nonlinearity, 26 (2013), 691–709. https://doi.org/10.1088/0951-7715/26/3/691 doi: 10.1088/0951-7715/26/3/691
|
| [15] |
D. Xiao, Dynamics and bifurcations on a class of population model with seasonal constant-yield harvesting, Discrete Contin. Dyn. Syst. B, 21 (2016), 699–719. https://doi.org/10.3934/dcdsb.2016.21.699 doi: 10.3934/dcdsb.2016.21.699
|
| [16] |
P. G. Barrientos, J. A. Rodriguez, A. Ruiz-Herrera, Chaotic dynamics in the seasonally forced SIR epidemic model, J. Math. Biol., 75 (2017), 1655–1668. https://doi.org/10.1007/s00285-017-1130-9 doi: 10.1007/s00285-017-1130-9
|
| [17] |
M. Han, X. Hou, L. Sheng, C. Wang, Theory of rotated equations and applications to a population model, Discrete Contin. Dyn. Syst., 38 (2018), 2171–2185. https://doi.org/10.3934/dcds.2018089 doi: 10.3934/dcds.2018089
|
| [18] |
Y. Tang, D. Xiao, W. Zhang, D. Zhu, Dynamics of epidemic models with asymptomatic infection and seasonal succession, Math. Biosci. Eng., 14 (2017), 1407–1424. https://doi.org/10.3934/mbe.2017073 doi: 10.3934/mbe.2017073
|
| [19] |
H. Amann, Fixed point equations and nonlinear eigenvalue problems in Ordered Banach Spaces, SIAM Rev., 18 (1976), 620–709. https://doi.org/10.1137/1018114 doi: 10.1137/1018114
|
| [20] |
X. Liang, J. Jiang, On the finite-dimensional dynamical systems with limited competition, Trans. Am. Math. Soc., 354 (2002), 3535–3554. https://doi.org/10.1090/S0002-9947-02-03032-5 doi: 10.1090/S0002-9947-02-03032-5
|
| [21] |
E. C. Balreira, S. Elaydi, R. Luís, Global stability of higher dimensional monotone maps, J. Differ. Equation Appl., 23 (2017), 2037–2071. https://doi.org/10.1080/10236198.2017.1388375 doi: 10.1080/10236198.2017.1388375
|
| [22] |
A. Ruiz-Herrera, Exclusion and dominance in discrete population models via the carrying simplex, J. Differ. Equation Appl., 19 (2013), 96–113. https://doi.org/10.1080/10236198.2011.628663 doi: 10.1080/10236198.2011.628663
|
| [23] |
A. Ruiz-Herrera, Topological criteria of global attraction with applications in population dynamics, Nonlinearity, 25 (2012), 2823–2841. https://doi.org/10.1088/0951-7715/25/10/2823 doi: 10.1088/0951-7715/25/10/2823
|
| [24] |
H. L. Smith, Competing subcommunities of mutualists and a generalized Kamke theorem, SIAM J. Math. Anal., 46 (1986), 856–874. https://doi.org/10.1137/0146052 doi: 10.1137/0146052
|
| [25] | H. L. Smith, P. Waltman, The Theory of the Chemostat, Cambridge University Press, 1995. https://doi.org/10.1017/CBO9780511530043 |
| [26] |
Y. Wang, J. Jiang, Uniqueness and attractivity of the carrying simplex for discrete-time competitive dynamical systems, J. Differ. Equations, 186 (2002), 611–632. https://doi.org/10.1016/S0022-0396(02)00025-6 doi: 10.1016/S0022-0396(02)00025-6
|




Figures(5)

Xizhuang Xie, Meixiang Chen. Global stability of a tridiagonal competition model with seasonal succession[J]. Mathematical Biosciences and Engineering, 2023, 20(4): 6062-6083. doi: 10.3934/mbe.2023262







 DownLoad:
DownLoad: