Citation: Lale Asik, Jackson Kulik, Kevin Long, Angela Peace. Dynamics of a stoichiometric producer-grazer system with seasonal effects on light level[J]. Mathematical Biosciences and Engineering, 2019, 16(1): 501-515. doi: 10.3934/mbe.2019023

| [1] | R.W. Sterner, and J.J. Elser, Ecological stoichiometry: the biology of elements from molecules to the biosphere, Princeton University Press, 2002. |
| [2] | M. Danger and F. Maunoury-Danger, Ecological Stoichiometry, in Encyclopedia of Aquatic Ecotoxicology (eds. J.F. Frard, and B. Christian), Springer, (2013), 317–326. |
| [3] | V.H. Smith, Nutrient dependence of primary productivity in lakes, Limnol. Oceanogr., 24 (1979), 1051-1064. |
| [4] | I. Loladze, Y. Kuang and J.J. Elser, Stoichiometry in producer-grazer systems: Linking energy flow with element cycling, Bull. Math Biol., 62 (2000), 1137-1162. |
| [5] | S.E. Jørgensen and B.D. Fath, Modelling Population Dynamics, in Developments in Environmental Modelling, Elsevier, (2011), 129–158. |
| [6] | S.E. Jorgenson and G. Bendoricchio, Fundamentals of ecological modelling, Elsevier Science Publishers BV, 2001. |
| [7] | T. Andersen, Pelagic Nutrient Cycles: Herbivores as Sources and Sinks, NY: Springer-Verlag, 1997. |
| [8] | J. Urabe and R.W. Sterner, Regulation of herbivore growth by the balance of light and nutrients, Proc. Nat. Acad. Sci, 93 (1996), 8465–8469. |
| [9] | J. Cebrián and I.I. Valiela, Seasonal patterns in phytoplankton biomass in coastal ecosystems, J. Plankton Res., 21 (1999), 429–444. |
| [10] | M.L. Rosenzweig, Paradox of enrichment: destabilization of exploitation ecosystems in ecological time, Science, 171 (1971), 385–387. |
| [11] | S. Lynch, Maps and Nonautonomous Systems in the Plane, in Dynamical Systems with Applications using Mathematica®, Birkhüser Boston, (2007), 171--194. |
| [12] | M. Scheffer, S. Rinaldi, Y.A. Kuznetsov and E.H. van Nes, Seasonal dynamics of Daphnia and algae explained as a periodically forced predator-prey system, Oikos, (1997), 519–532. |
| [13] | S. Ulrich, G.Z. Maciej, L. Winfried and D. Annie, The PEG-model of seasonal succession of planktonic events in fresh waters, Arch. Hydrobiol, 106 (1986), 433–471. |
| [14] | H. Wang, Y. Kuang and I. Loladze, Dynamics of a mechanistically derived stoichiometric producer-grazer model, J. Biol. Dynam., 2 (2008), 286–296. |
| [15] | I. Loladze, Y. Kuang, J.J. Elser and W.F. Fagan, Competition and stoichiometry: coexistence of two predators on one prey, Theor. Popul. Biol., 65 (2004), 1–15. |
| [16] | J. Urabe, J.J. Elser, M. Kyle, T. Yoshida, T. Sekino and Z. Kawabata, Herbivorous animals can mitigate unfavourable ratios of energy and material supplies by enhancing nutrient recycling, Ecol. Lett., 5 (2002), 177–185. |
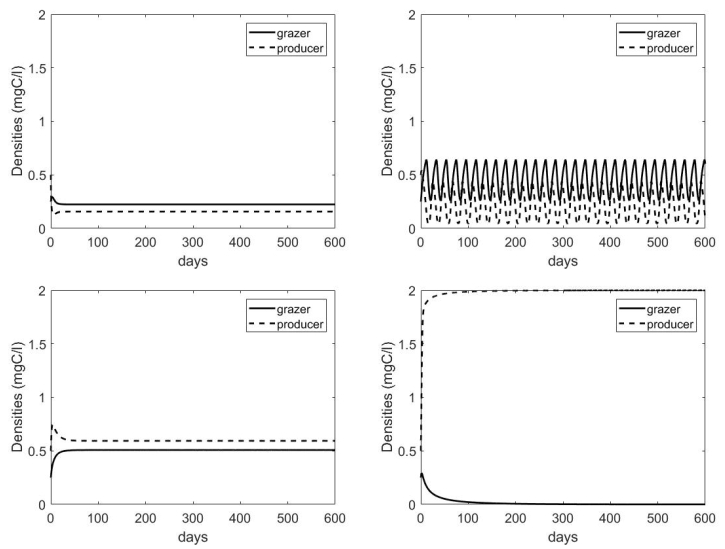
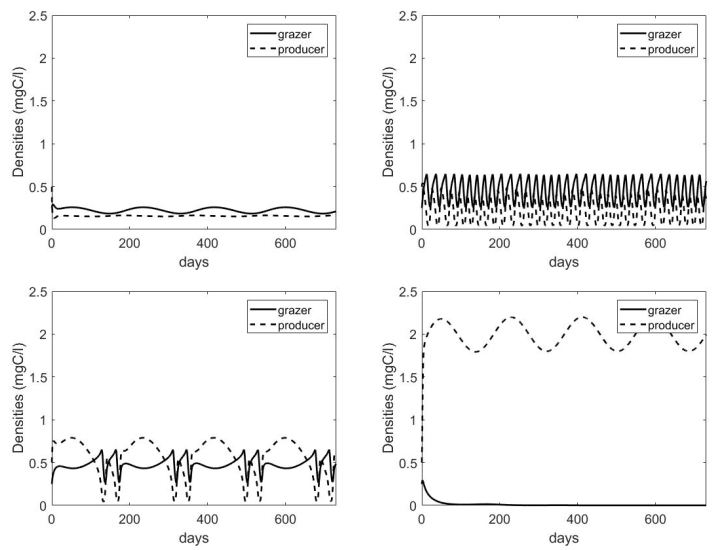
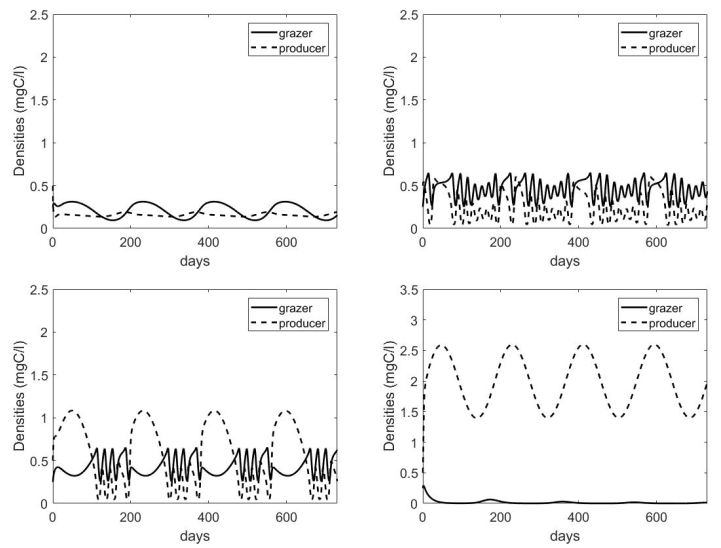
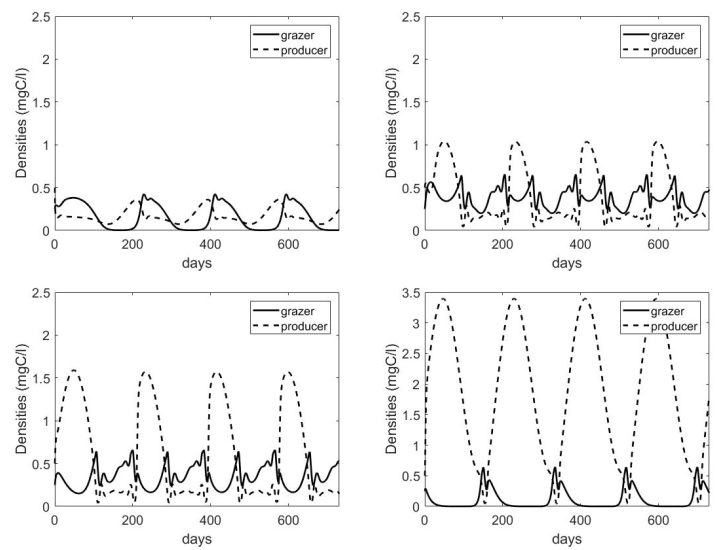
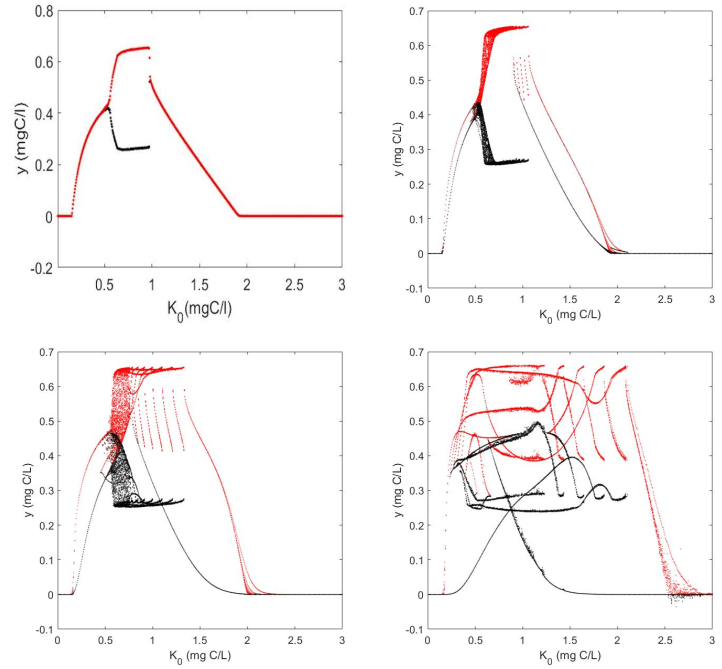
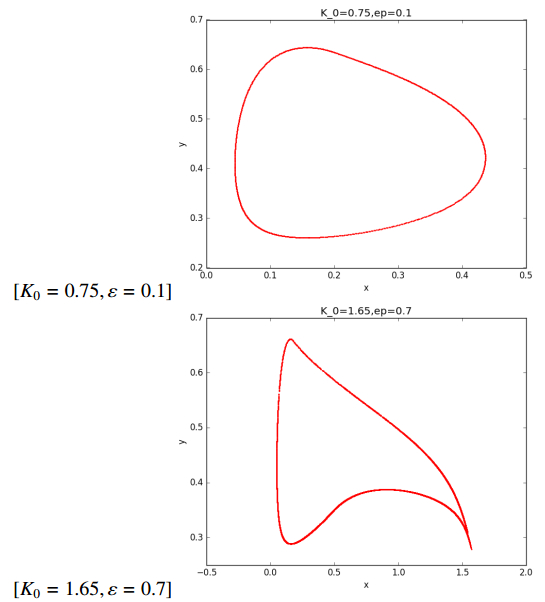
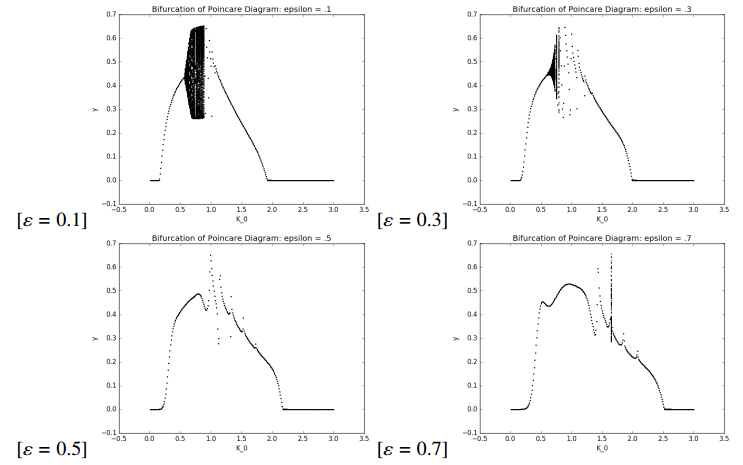
Figures(7) / Tables(1)

Lale Asik, Jackson Kulik, Kevin Long, Angela Peace. Dynamics of a stoichiometric producer-grazer system with seasonal effects on light level[J]. Mathematical Biosciences and Engineering, 2019, 16(1): 501-515. doi: 10.3934/mbe.2019023







 DownLoad:
DownLoad: