The population of biological species in the ecosystem is known sensitive to the periodic fluctuations of seasonal change, food resources and climatic conditions. Research in the ecological management discipline conventionally models the behavior of such dynamic systems through specific impulsive response functions, but the results of such research are applicable only when the environments conform exactly to the conditions as defined by the specific response functions that have been implemented for specific scenarios. This means that the application of previous work may be somewhat limited. Moreover, the intra and inter competitions among species have been seldom studied for modelling the prey-predator ecosystem. To fill in the gaps this paper models the delicate balance of two-prey and one-predator system by addressing three main areas of: ⅰ) instead of using the specific impulse response this work models the ecosystem through a more general response function; ⅱ) to include the effects due to the competition between species and ⅲ) the system is subjected to the influences of seasonal factors. The seasonal factor has been implemented here in terms of periodic functions to represent the growth rates of predators. The sufficient condition for the local and global asymptotic stability of the prey-free periodic solution and the permanence of the system have been subsequently obtained by using the Comparison techniques and the Floquet theorems. Finally, the correctness of developed theories is verified by numerical simulation, and the corresponding biological explanation is given.
Citation: Jie Hu, Juan Liu, Peter Yuen, Fuzhong Li, Linqiang Deng. Modelling of a seasonally perturbed competitive three species impulsive system[J]. Mathematical Biosciences and Engineering, 2022, 19(3): 3223-3241. doi: 10.3934/mbe.2022149
The population of biological species in the ecosystem is known sensitive to the periodic fluctuations of seasonal change, food resources and climatic conditions. Research in the ecological management discipline conventionally models the behavior of such dynamic systems through specific impulsive response functions, but the results of such research are applicable only when the environments conform exactly to the conditions as defined by the specific response functions that have been implemented for specific scenarios. This means that the application of previous work may be somewhat limited. Moreover, the intra and inter competitions among species have been seldom studied for modelling the prey-predator ecosystem. To fill in the gaps this paper models the delicate balance of two-prey and one-predator system by addressing three main areas of: ⅰ) instead of using the specific impulse response this work models the ecosystem through a more general response function; ⅱ) to include the effects due to the competition between species and ⅲ) the system is subjected to the influences of seasonal factors. The seasonal factor has been implemented here in terms of periodic functions to represent the growth rates of predators. The sufficient condition for the local and global asymptotic stability of the prey-free periodic solution and the permanence of the system have been subsequently obtained by using the Comparison techniques and the Floquet theorems. Finally, the correctness of developed theories is verified by numerical simulation, and the corresponding biological explanation is given.

| [1] |
R. Tyson, F. Lutscher, Seasonally varying predation behavior and climate shifts are predicted to affect predator-prey cycles, Am. Nat., 188 (2016), 539–553. https://doi.org/10.1086/688665 doi: 10.1086/688665

|
| [2] |
X. H. Wang, C. Y. Huang, Permanence of a stage-structured predator-prey system with impulsive stocking prey and harvesting predator, Appl. Math. Comput., 235 (2014), 32–42. https://doi.org/10.1016/j.amc.2014.02.092 doi: 10.1016/j.amc.2014.02.092
|
| [3] |
H. Zhu, P. Li, X. Li, H. Akca, Input-to-state stability for impulsive switched systems with incommensurate impulsive switching signals, Commun. Nonlinear. Sci., 80 (2020), 104969. https://doi.org/10.1016/j.cnsns.2019.104969 doi: 10.1016/j.cnsns.2019.104969
|
| [4] |
Z. Wang, Y. Xie, J. Lu, Y. Li, Stability and bifurcation of a delayed generalized fractional-order prey-predator model with interspecific competition, Appl. Math. Comput., 347 (2019), 360–369. https://doi.org/10.1016/j.amc.2018.11.016 doi: 10.1016/j.amc.2018.11.016
|
| [5] |
X. Wang, Z. Wang, H. Shen, Dynamical analysis of a discrete-time SIS epidemic model on complex networks, Appl. Math. Lett., 94 (2019), 292–299. https://doi.org/10.1016/j.aml.2019.03.011 doi: 10.1016/j.aml.2019.03.011
|
| [6] |
G. R. Liu, Z. M. Liu, Z. Jin, analysis of epidemic and information spreading in overlay networks, J. Theor. Biol., 444 (2018), 28–37. https://doi.org/10.1016/j.jtbi.2018.02.010 doi: 10.1016/j.jtbi.2018.02.010
|
| [7] |
Z. Wang, Y. Shao, X. Fang, The dynamic behaviors of one-predator two prey system with mutual interference and impulsive control, Math. Comput. Simulat., 132 (2017), 68–65. https://doi.org/10.1016/j.matcom.2016.06.007 doi: 10.1016/j.matcom.2016.06.007
|
| [8] |
W. Li, J. Ji, L. Huang, Global dynamic behavior of a predator-prey model under ratio-dependent state impulsive control, Appl. Math. Model., 77 (2020), 1842–1859. https://doi.org/10.1016/j.apm.2019.09.033 doi: 10.1016/j.apm.2019.09.033
|
| [9] |
Z. J. Du, Z. S. Feng, Periodic solutions of a neutral impulsive predator-prey model with Beddington-DeAngelis functional response with delays, Appl. Math. Comput., 258 (2014), 87–98. https://doi.org/10.1016/j.cam.2013.09.008 doi: 10.1016/j.cam.2013.09.008
|
| [10] |
R. Kaushik, S. Banerjee, Predator-prey system: Prey's counter-attack on juvenile predators shows opposite side of the same ecological coin, Appl. Math. Comput., 388 (2021), 125530. https://doi.org/10.1016/j.amc.2020.125530 doi: 10.1016/j.amc.2020.125530
|
| [11] |
S. Khajanchi, S. Banerjee, Role of constant prey refuge on stage structure predator-prey model with ratio dependent functional response, Appl. Math. Comput., 314 (2017), 193–198. https://doi.org/10.1016/j.amc.2017.07.017 doi: 10.1016/j.amc.2017.07.017
|
| [12] |
Z. J. Liu, S. M. Zhong, Z. D. Teng, L. Zhang, Permanence and global attractivity of an impulsive ratio-dependent predator-prey system in a patchy environment, Appl. Math. Comput., 219 (2013), 9791–9804. https://doi.org/10.1016/j.amc.2013.03.108 doi: 10.1016/j.amc.2013.03.108
|
| [13] |
C.V. Kumar, K. S. Reddy, M. Srinivas, Dynamics of prey predator with Holling interactions and stochastic influences, Alex. Eng. J., 57(2018), 1079–1086. https://doi.org/10.1016/j.aej.2017.02.004 doi: 10.1016/j.aej.2017.02.004
|
| [14] |
A. Izquierdo, M. Len, M. Mayado, A generalized Holling type Ⅱ model for the interaction between dextral-sinistral snails and Pareas snakes, Appl. Math. Model., 73 (2019), 459–472. https://doi.org/10.1016/j.apm.2019.04.005 doi: 10.1016/j.apm.2019.04.005
|
| [15] | H. Liu, Dynamic analysis of impulsively controlled predator-prey model with general functional response and seasonal effect, International Conference on Artificial Intelligence and Computational Intelligence. IEEE Computer Society, (2010), 322–330. |
| [16] |
X. Wang, J. Wei, Dynamics in a diffusive predator-prey system with strong Allee effect and Ivlev-type functional response, J. Math. Anal. Appl., 422 (2015), 1447–1462. https://doi.org/10.1016/j.jmaa.2014.09.051 doi: 10.1016/j.jmaa.2014.09.051
|
| [17] |
H. Kim, H. Baek, The dynamical complexity of a predator-prey system with Hassell-Varley functional response and impulsive effect, Math. Comput. Simulat., 9 (2013), 1–14. https://doi.org/10.1016/j.matcom.2013.05.011 doi: 10.1016/j.matcom.2013.05.011
|
| [18] |
Z. Yao, S. Xie, N. Yu, Dynamics of cooperative predator-prey system with impulsive effects and Beddington-DeAngelis functional response, J. Egypt. Math. Soc., 21 (2013), 213–223. https://doi.org/10.1016/j.joems.2013.04.008 doi: 10.1016/j.joems.2013.04.008
|
| [19] |
J. Liu, J. Hu, P. Yuen, Extinction and permanence of the predator-prey system with general functional response and impulsive control, Appl. Math. Model., 88 (2020), 55–67. https://doi.org/10.1016/j.apm.2020.06.033 doi: 10.1016/j.apm.2020.06.033
|
| [20] |
C. Lu, X. H. Ding, Periodic solutions and stationary distribution for a stochastic predator-prey system with impulsive perturbations, Appl. Math. Comput., 350 (2019), 313–322. https://doi.org/10.1016/j.amc.2019.01.023 doi: 10.1016/j.amc.2019.01.023
|
| [21] |
Y. X. Xie, L. J. Wang, Q. C. Deng, Z. J. Wu, The dynamics of an impulsive predator-prey model with communicable disease in the prey species only, Appl. Math. Comput., 292 (2017), 320–335. https://doi.org/10.1016/j.amc.2016.07.042 doi: 10.1016/j.amc.2016.07.042
|
| [22] |
X. H. Wang, C.Y. Huang, Permanence of a stage-structured predator-prey system with impulsive stocking prey and harvesting predator, Appl. Math. Comput., 235 (2014), 32–42. https://doi.org/10.1016/j.amc.2014.02.092 doi: 10.1016/j.amc.2014.02.092
|
| [23] |
X. W. Jiang, Q. Song, M.Y. Hao, Dynamics behaviors of a delayed stage-structured predator-prey model with impulsive effect, Appl. Math. Comput., 215 (2010), 4221–4229. https://doi.org/10.1016/j.amc.2009.12.044 doi: 10.1016/j.amc.2009.12.044
|
| [24] |
H. D. Cheng, T. Q. Zhang, A new predator-prey model with a profitless delay of digestion and impulsive perturbation on the prey, Appl. Math. Comput., 217 (2011), 9198–9208. https://doi.org/10.1016/j.amc.2011.03.159 doi: 10.1016/j.amc.2011.03.159
|
| [25] |
Z. Zhao, X. B. Wu, Nonlinear analysis of a delayed stage-structured predator-prey model with impulsive effect and environment pollution, Appl. Math. Comput., 232 (2014), 1262–1268. https://doi.org/10.1016/j.amc.2014.01.003 doi: 10.1016/j.amc.2014.01.003
|
| [26] |
Y. Z. Pei, C. G. Li, S. H. Fan, A mathematical model of a three species prey-predator system with impulsive control and Holling functional response, Appl. Math. Comput., 219 (2013), 10945–10955. https://doi.org/10.1016/j.amc.2013.05.012 doi: 10.1016/j.amc.2013.05.012
|
| [27] |
S. Li, W. Liu, A delayed Holling type Ⅲ functional response predator prey system with impulsive perturbation on the prey, Adv. Differ. Equ-ny., 42 (2016), 42–62. https://doi.org/10.1186/s13662-016-0768-8 doi: 10.1186/s13662-016-0768-8
|
| [28] |
X. Wang, J. Wei, Dynamics in a diffusive predator-prey system with strong Allee effect and Ivlev-type functional response, J. Math. Anal. Appl., 422 (2015), 1447–1462. https://doi.org/10.1016/j.jmaa.2014.09.051 doi: 10.1016/j.jmaa.2014.09.051
|
| [29] |
L. Hang, L. Zhang, X. W. Wang, A hybrid predator-prey model with general functional responses under seasonal succession alternating between Gompertz and logistic growth, Adv. Differ. Equ-ny., 10 (2020). https://doi.org/10.1186/s13662-019-2477-6 doi: 10.1186/s13662-019-2477-6
|
| [30] |
G. R. Liu, J. R. Yan, Global asymptotic stability of nonlinear neutral differential equation, Commun. Nonlinear. Sci., 19 (2014), 1035–1041. https://doi.org/10.1016/j.cnsns.2013.08.035 doi: 10.1016/j.cnsns.2013.08.035
|
| [31] | V. Lakshmikantham, D. Bainov, P. Simeonov, Theory of Impulsive Differential Equations, World Scientific Publisher, Singapore, (1989), 108–190. |
| [32] |
Z. J. Wang, M. liu, Optimal impulsive harvesting strategy of a stochastic Gompertz model in periodic environments, Appl. Math. Lett., 125(2022), 107733. https://doi.org/10.1016/j.aml.2021.107733 doi: 10.1016/j.aml.2021.107733
|
| [33] |
S. Q. Zhang, S. L. Yuan, T. H. Zhang, A predator-prey model with different response functions to juvenile and adult prey in deterministic and stochastic environments, Appl. Math. Comput., 413 (2022), 126598. https://doi.org/10.1016/j.amc.2021.126598 doi: 10.1016/j.amc.2021.126598
|
| [34] |
C. Lu, Dynamical analysis and numerical simulations on a crowley-Martin predator-prey model in stochastic environment, Appl. Math. Comput., 413 (2022), 126641. https://doi.org/10.1016/j.amc.2021.126641 doi: 10.1016/j.amc.2021.126641
|
| [35] |
X. H. Zhang, Q. Yang, Threshold behavior in a stochastic SVIR model with general incidence rates, Appl. Math. Lett., 121 (2021), 107403. https://doi.org/10.1016/j.aml.2021.107403 doi: 10.1016/j.aml.2021.107403
|
| [36] |
M. liu, C. Z. Bai, Optimal harvesting of a stochastic mutualism model with regime-switching, Appl. Math. Comput., 373 (2020), 125040. https://doi.org/10.1016/j.amc.2020.125040 doi: 10.1016/j.amc.2020.125040
|
| [37] |
C. Lu, X. H. Ding, Periodic solutions and stationary distribution for a stochastic predator-prey system with impulsive perturbations, Appl. Math. Comput., 350 (2019), 313–322. https://doi.org/10.1016/j.amc.2019.01.023 doi: 10.1016/j.amc.2019.01.023
|
| [38] |
H. K. Qi, X. Z. Meng, Mathematical modeling, analysis and numerical simulation of HIV: The influence of stochastic environmental fluctuations on dynamics, Math. Comput. Simulat., 187 (2021), 700–719. https://doi.org/10.1016/j.matcom.2021.03.027 doi: 10.1016/j.matcom.2021.03.027
|
| [39] |
H. K. Qi, X. Z. Meng, T. Hayat, A. Hobiny, Stationary distribution of a stochastic predator-prey model with hunting cooperation, Appl. Math. Lett., 124 (2022), 107662. https://doi.org/10.1016/j.aml.2021.107662 doi: 10.1016/j.aml.2021.107662
|
| [40] |
H. K. Qi, X. Z. Meng, Threshold behavior of a stochastic predator-prey system with prey refuge and fear effect, Appl. Math. Lett., 113 (2021), 106846. https://doi.org/10.1016/j.aml.2020.106846 doi: 10.1016/j.aml.2020.106846
|
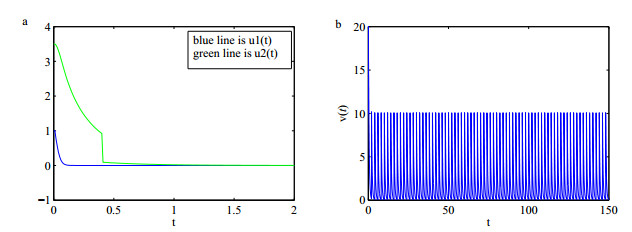
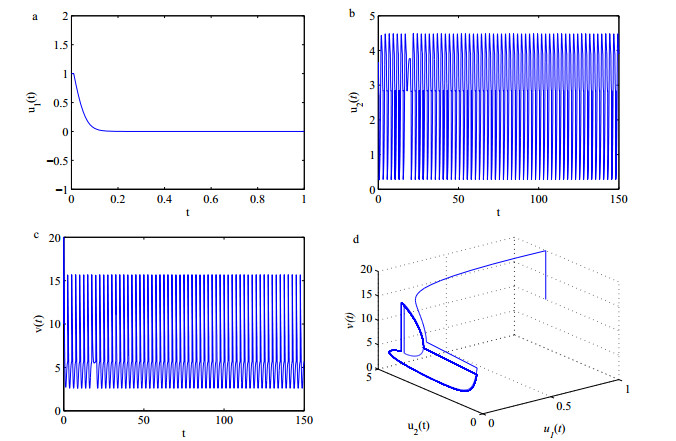
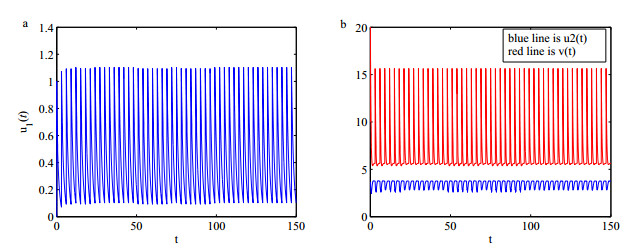
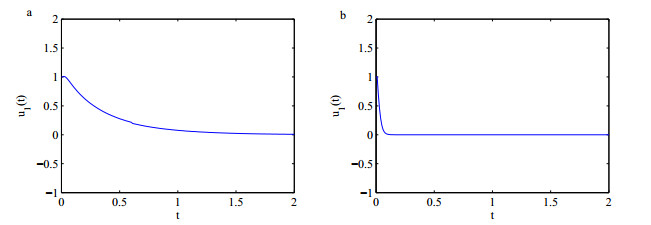
Figures(5)

Jie Hu, Juan Liu, Peter Yuen, Fuzhong Li, Linqiang Deng. Modelling of a seasonally perturbed competitive three species impulsive system[J]. Mathematical Biosciences and Engineering, 2022, 19(3): 3223-3241. doi: 10.3934/mbe.2022149







 DownLoad:
DownLoad: