In this paper, a new stochastic predator-prey model with impulsive perturbation and Crowley-Martin functional response is proposed. The dynamical properties of the model are systematically investigated. The existence and stochastically ultimate boundedness of a global positive solution are derived using the theory of impulsive stochastic differential equations. Some sufficient criteria are obtained to guarantee the extinction and a series of persistence in the mean of the system. Moreover, we provide conditions for the stochastic permanence and global attractivity of the model. Numerical simulations are performed to support our qualitative results.
Citation: Yan Zhang, Shujing Gao, Shihua Chen. Modelling and analysis of a stochastic nonautonomous predator-prey model with impulsive effects and nonlinear functional response[J]. Mathematical Biosciences and Engineering, 2021, 18(2): 1485-1512. doi: 10.3934/mbe.2021077
In this paper, a new stochastic predator-prey model with impulsive perturbation and Crowley-Martin functional response is proposed. The dynamical properties of the model are systematically investigated. The existence and stochastically ultimate boundedness of a global positive solution are derived using the theory of impulsive stochastic differential equations. Some sufficient criteria are obtained to guarantee the extinction and a series of persistence in the mean of the system. Moreover, we provide conditions for the stochastic permanence and global attractivity of the model. Numerical simulations are performed to support our qualitative results.

| [1] | H. I. Freedman, Deterministic Mathematical Models in Population Ecology, Marcel Dekker, New York, 1980. |
| [2] |
Z. Yao, S. Xie, N. Yu, Dynamics of cooperative predator-prey system with impulsive effects and Beddington-DeAngelis functional response, J. Egypt. Math. Soc., 21 (2013), 213-223. doi: 10.1016/j.joems.2013.04.008

|
| [3] |
Z. Shen, J. Wei, Hopf bifurcation analysis in a diffusive predator-prey system with delay and surplus killing effect, Math. Biosci. Eng., 15 (2018), 693-715. doi: 10.3934/mbe.2018031
|
| [4] |
M. Bandyopadhyay, J. Chattopadhyay, Ratio-dependent predator-prey model: Effect of environmental fluctuation and stability, Nonlinearity, 18 (2005), 913-936. doi: 10.1088/0951-7715/18/2/022
|
| [5] |
M. Liu, K. Wang, Q. Wu, Survival analysis of stochastic competitive models in a polluted environment and stochastic competitive exclusion principle, B. Math. Biol., 73 (2011), 1969-2012. doi: 10.1007/s11538-010-9569-5
|
| [6] |
M. Liu, C. Bai, K. Wang, Asymptotic stability of a two-group stochastic SEIR model with infinite delays, Commun. Nonlinear. Sci. Numer. Simulat., 19 (2014), 3444-3453. doi: 10.1016/j.cnsns.2014.02.025
|
| [7] |
X. Mao, G. Marion, E. Renshaw, Environmental Brownian noise suppresses explosions in populations dynamics, Stoch. Proc. Appl., 97 (2002), 95-110. doi: 10.1016/S0304-4149(01)00126-0
|
| [8] | D. Jana, R. Agrawal, R. K. Upadhyay, Dynamics of generalist predator in a stochastic environment: Effect of delayed growth and prey refuge, Appl. Math. Comput., 268 (2015), 1072-1094. |
| [9] | Y. Lin, D. Jiang, S. Wang, Stationary distribution of a stochastic SIS epidemic model with vaccination, Physica A, 394 (2014), 718-727. |
| [10] |
M. Liu, K. Wang, Persistence and extinction in stochastic non-autonomous logistic systems, J. Math. Anal. Appl., 375 (2011), 443-457. doi: 10.1016/j.jmaa.2010.09.058
|
| [11] |
M. Liu, K. Wang, Persistence, extinction and global asymptotical stability of a non-autonomous predator-prey model with random perturbation, Appl. Math. Model., 36 (2012), 5344-5353. doi: 10.1016/j.apm.2011.12.057
|
| [12] |
Y. Zhao, S. Yuan, J. Ma, Survival and stationary distribution analysis of a stochastic competitive model of three species in a polluted environment, B. Math. Biol., 77 (2015), 1285-1326. doi: 10.1007/s11538-015-0086-4
|
| [13] |
L. Zu, D. Jiang, D. O'Regan, B. Ge, Periodic solution for a non-autonomous Lotka-Volterra predator-prey model with random perturbation, J. Math. Anal. Appl., 430 (2015), 428-437. doi: 10.1016/j.jmaa.2015.04.058
|
| [14] |
X. Liu, S. Zhong, B. Tian, F. Zheng, Asymptotic properties of a stochastic predator-prey model with Crowley-Martin functional response, J. Appl. Math. Comput., 43 (2013), 479-490. doi: 10.1007/s12190-013-0674-0
|
| [15] |
M. Hassell, G. Varley, New inductive population model for insect parasites and its bearing on biologicalcontrol, Nature, 223 (1969), 1133-1137. doi: 10.1038/2231133a0
|
| [16] |
X. Yan, C. Zhang, Stability and turing instability in a diffusive predator-prey system with Beddington-DeAngelis functional response, Nonlinear Anal-Real., 20 (2014), 1-13. doi: 10.1016/j.nonrwa.2014.04.001
|
| [17] |
C. Xu, P. Li, 20 Oscillations for a delayed predator-prey model with Hassell-Varley-type functional response, C. R. Biol., 338 (2015), 227-240. doi: 10.1016/j.crvi.2015.01.002
|
| [18] |
Y. Zhang, S. Gao, Y. Liu, Analysis of a nonautonomous model for migratory birds with saturation incidence rate, Commun. Nonlinear Sci. Numer. Simulat., 17 (2012), 1659-1672. doi: 10.1016/j.cnsns.2011.08.040
|
| [19] | R. May, Stability and Complexity in Model Ecosystems, Princeton: Princeton University Press, 1974. |
| [20] |
X. Shi, X. Zhou, X. Song, Analysis of a stage-structured predator-prey model with Crowley-Martin function, J. Appl. Math. Comput., 36 (2011), 459-472. doi: 10.1007/s12190-010-0413-8
|
| [21] |
T. Zhang, J. Zhang, X. Meng, Geometric analysis of a pest management model with Holling's type III functional response and nonlinear state feedback control, Nonlinear Dyn., 84 (2016), 1529-1539. doi: 10.1007/s11071-015-2586-z
|
| [22] |
S. Chen, J. Wei, J. Yu, Stationary patterns of a diffusive predator-prey model with Crowley-Martin functional response, Nonlinear Anal. Real, 39 (2018), 33-57. doi: 10.1016/j.nonrwa.2017.05.005
|
| [23] |
J. Tripathi, S. Tyagi, S. Abbas, Global analysis of a delayed density dependent predator Cprey model with Crowley-Martin functional response, Commun. Nonlinear. Sci. Numer. Simulat., 30 (2016), 45-69. doi: 10.1016/j.cnsns.2015.06.008
|
| [24] | R. Tan, Z. Liu, S. Guo, H. Xiang, On a nonautonomous competitive system subject to stochastic and impulsive perturbations, Appl. Math. Comput., 256 (2015), 702-714. |
| [25] |
S. Zhang, D. Tan, Dynamics of a stochastic predator-prey system in a polluted environment with pulse toxicant input and impulsive perturbations, Appl. Math. Model., 39 (2015), 6319-6331. doi: 10.1016/j.apm.2014.12.020
|
| [26] |
R. Wu, X. Zou, K. Wang, Asymptotic behavior of a stochastic non-autonomous predator-prey model with impulsive perturbations, Commun. Nonlinear Sci. Numer. Simulat., 20 (2015), 965-974. doi: 10.1016/j.cnsns.2014.06.023
|
| [27] |
Y. Zhang, K. Fan, S. Gao, S. Chen, A remark on stationary distribution of a stochastic SIR epidemic model with double saturated rates, Appl. Math. lett., 76 (2018), 46-52. doi: 10.1016/j.aml.2017.08.002
|
| [28] |
X. Li, X. Mao, Population dynamical behavior of non-autonomous Lotka-Volterra competitive system with random perturbation, Discret Contin. Dyn. Syst., 24 (2009), 523-593. doi: 10.3934/dcds.2009.24.523
|
| [29] |
M. Liu and K. Wang, On a stochastic logistic equation with impulsive perturbations, Comput. Math. Appl., 63 (2012), 871-886. doi: 10.1016/j.camwa.2011.11.003
|
| [30] |
X. Mao, Stochastic versions of the Lassalle Theorem, T. Differ. Equ., 153 (1999), 175-195. doi: 10.1006/jdeq.1998.3552
|
| [31] | R. Tan, Z. Liu, R.A. Cheke, Periodicity and stability in a single-species model governed by impulsive differential equation, Appl. Math. Comput., 36 (2012), 1085-1094. |
| [32] |
M. Liu, M. Fan, Permanence of stochastic Lotka-Volterra systems, J. Nonlinear Sci., 27 (2017), 425-452. doi: 10.1007/s00332-016-9337-2
|
| [33] | K. Wang, Stochastic Models in Mathematical Biology, Beijing: Science Press, 2010. |
| [34] |
S. Cheng, Stochastic population systems, Stoch. Proc. Appl., 27 (2009), 854-874. doi: 10.1080/07362990902844348
|
| [35] |
D. Higham, An algorithmic introduction to numerical simulation of stochastic differential equations, SIAM Rev., 43 (2001), 525-546. doi: 10.1137/S0036144500378302
|
| [36] | M. Liu, C. Bai, Optimal harvesting of a stochastic mutualism model with regime-switching, Appl. Math. Comput., 373 (2020). |
| [37] | W. Ji, Z. Wang, G. Hu, Stationary distribution of a stochastic hybrid phytoplankton model with allelopathy, Adv. Differ. Equ. NY, 2020. |
| [38] | Z. Wang, M. Deng, M. Liu, Stationary distribution of a stochastic ratio-dependent predator-prey system with regime-switching, Chaos Soliton Fract., 2020, 110462. |
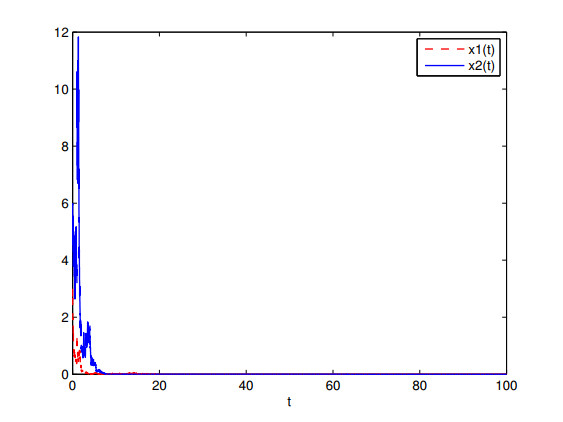
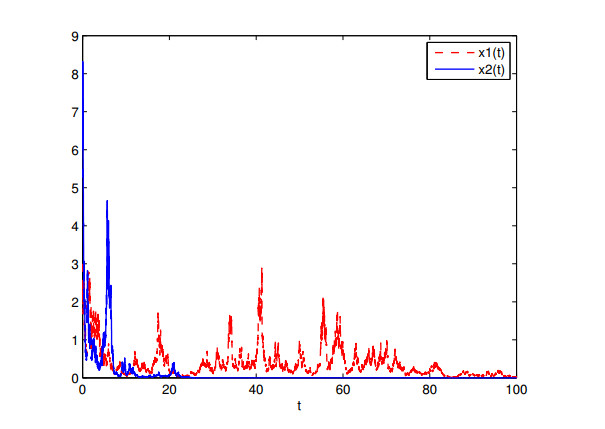
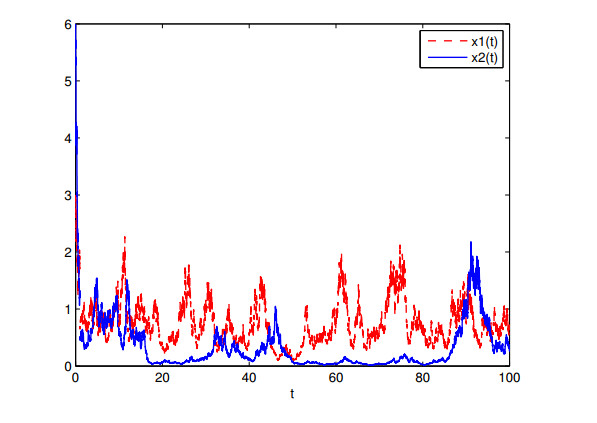
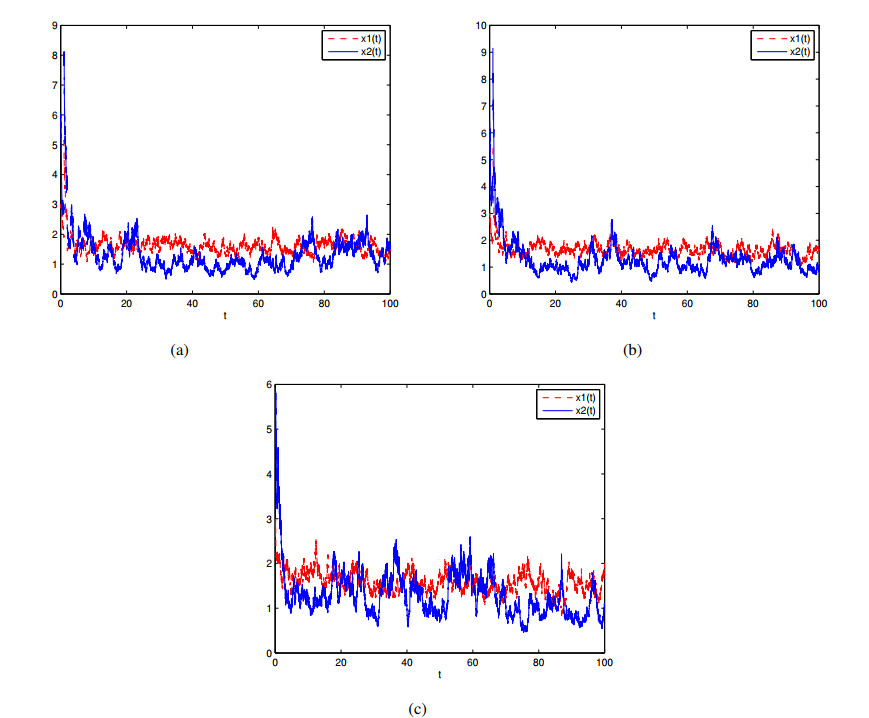
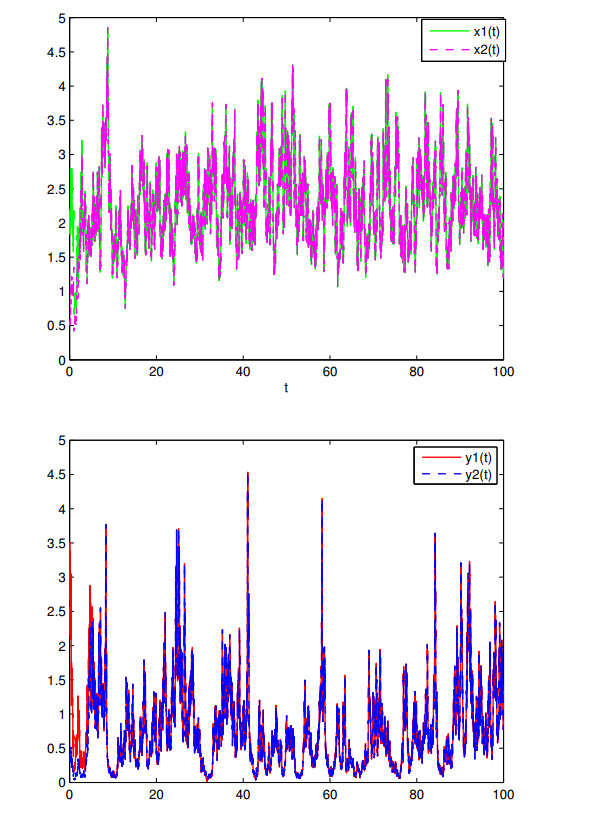
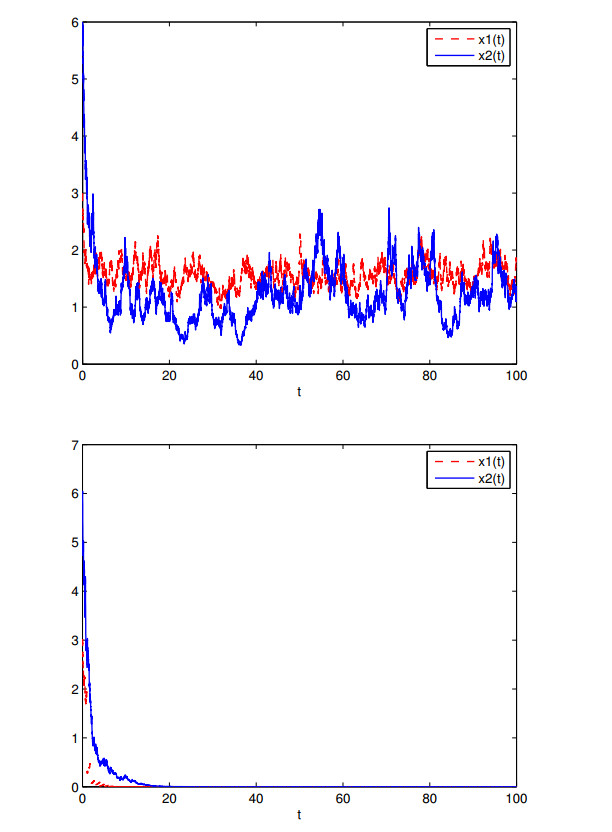
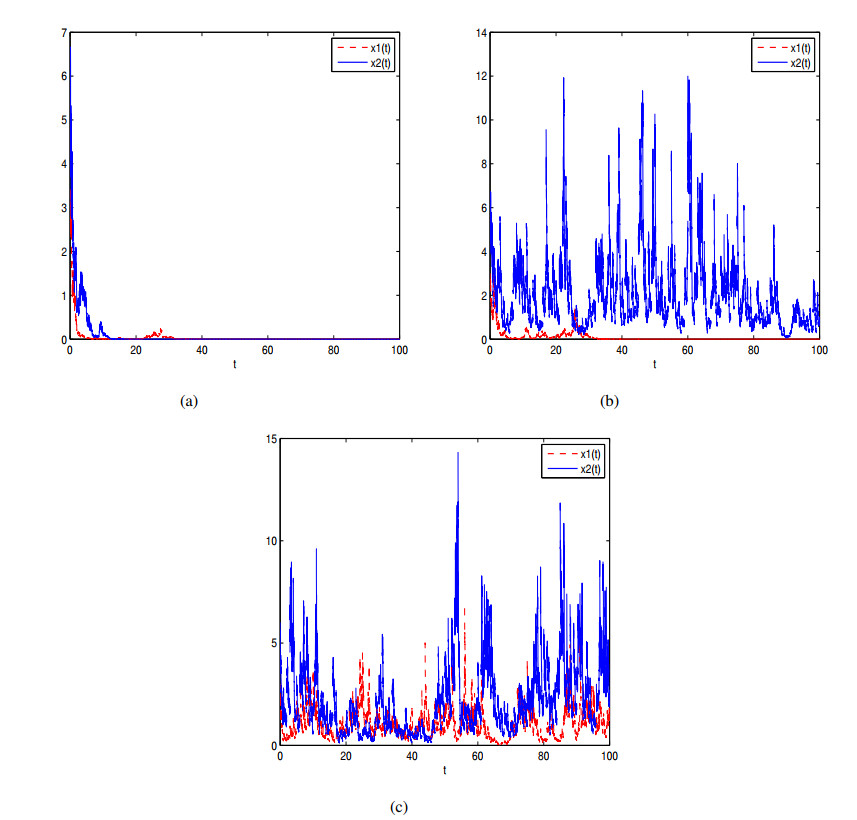
Figures(7)

Yan Zhang, Shujing Gao, Shihua Chen. Modelling and analysis of a stochastic nonautonomous predator-prey model with impulsive effects and nonlinear functional response[J]. Mathematical Biosciences and Engineering, 2021, 18(2): 1485-1512. doi: 10.3934/mbe.2021077







 DownLoad:
DownLoad: