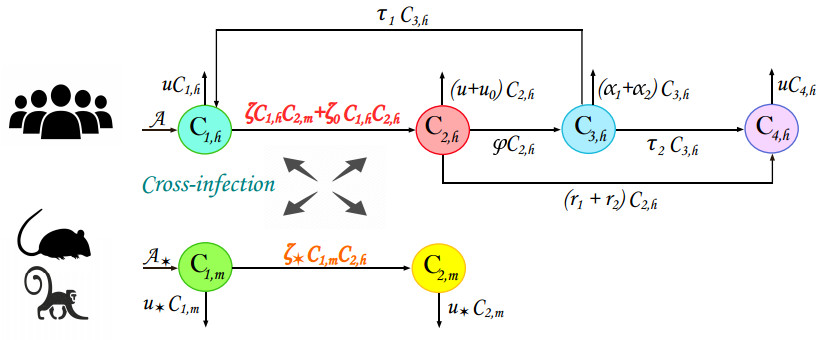
Monkeypox 2022, a new re-emerging disease, is caused by the Monkeypox virus. Structurally, this virus is related to the smallpox virus and infects the host in a similar way; however, the symptoms of Monkeypox are more severe. In this research work, a mathematical model for understanding the dynamics of Monkeypox 2022 is suggested that takes into account two modes of transmission: horizontal human dissemination and cross-infection between animals and humans. Due to lack of substantial knowledge about the virus diffusion and the effect of external perturbations, the model is extended to the probabilistic formulation with Lévy jumps. The proposed model is a two block compartmental system that requires the form of Itô-Lévy stochastic differential equations. Based on some assumptions and nonstandard analytical techniques, two principal asymptotic properties are proved: the eradication and continuation in the mean of Monkeypox 2022. The outcomes of the study reveals that the dynamical behavior of the proposed Monkeypox 2022 system is chiefly governed by some parameters that are precisely correlated with the noise intensities. To support the obtained theoretical finding, examples based on numerical simulations and real data are presented at the end of the study. The numerical simulations also exhibit the impact of the innovative adopted mathematical techniques on the findings of this work.
Citation: Asad Khan, Yassine Sabbar, Anwarud Din. Stochastic modeling of the Monkeypox 2022 epidemic with cross-infection hypothesis in a highly disturbed environment[J]. Mathematical Biosciences and Engineering, 2022, 19(12): 13560-13581. doi: 10.3934/mbe.2022633
Monkeypox 2022, a new re-emerging disease, is caused by the Monkeypox virus. Structurally, this virus is related to the smallpox virus and infects the host in a similar way; however, the symptoms of Monkeypox are more severe. In this research work, a mathematical model for understanding the dynamics of Monkeypox 2022 is suggested that takes into account two modes of transmission: horizontal human dissemination and cross-infection between animals and humans. Due to lack of substantial knowledge about the virus diffusion and the effect of external perturbations, the model is extended to the probabilistic formulation with Lévy jumps. The proposed model is a two block compartmental system that requires the form of Itô-Lévy stochastic differential equations. Based on some assumptions and nonstandard analytical techniques, two principal asymptotic properties are proved: the eradication and continuation in the mean of Monkeypox 2022. The outcomes of the study reveals that the dynamical behavior of the proposed Monkeypox 2022 system is chiefly governed by some parameters that are precisely correlated with the noise intensities. To support the obtained theoretical finding, examples based on numerical simulations and real data are presented at the end of the study. The numerical simulations also exhibit the impact of the innovative adopted mathematical techniques on the findings of this work.

| [1] | WHO, Monkeypox, 2022. Abstract from: https://www.who.int/news-room/fact-sheets/detail/monkeypox. |
| [2] | N. P. Singh, S. Sharma, G. Ghai, A. Singh, A systematic review on epidermology of human Monkeypox virus, Ann. Rom. Soc. Cell Biol., 25 (2021), 602–610. |
| [3] |
M. J. Oladoye, Monkeypox: A neglected viral zoonotic disease, Electron. J. Med. Edu. Technol., 14 (2021), 2108. https://doi.org/10.30935/ejmets/10911 doi: 10.30935/ejmets/10911

|
| [4] | S. Deresinski, A case of Monkeypox in a returned traveler, Infect. Dis. Alert, 41 (2022). |
| [5] | V. Costello, M. Sowash, A. Gaur, M. Cardis, H. Pasieka, G. Wortmann, et al., Imported Monkeypox from international traveler, Maryland, USA, 2021, Emerg. Infect. Dis., 28 (2022), 1002–1005. https://doi.org/10.3201%2Feid2805.220292 |
| [6] |
O. J. Peter, S. Kumar, N. Kumari, F. A. Oguntolu, K. Oshinubi, R. Musa, Transmission dynamics of Monkeypox virus: A mathematical modelling approach, Model. Earth Syst. Environ., 8 (2022), 3423–3434. https://doi.org/10.1007/s40808-021-01313-2 doi: 10.1007/s40808-021-01313-2
|
| [7] | WHO, Risk assessment: Monkeypox multi-country outbreak, 2022. Available from: https://www.ecdc.europa.eu/en/publications-data/risk-assessment-monkeypox-multi-country-outbreak. |
| [8] |
E. Petersen, A. Kantele, M. Koopmans, D. A. A. Yinka-Ogunleye, C. Ihekweazu, A. Zumla, Human Monkeypox: Epidemiologic and clinical characteristics, diagnosis, and prevention, Infect. Dis. Clin., 33 (2019), 1027–1043. https://doi.org/10.1016/j.idc.2019.03.001 doi: 10.1016/j.idc.2019.03.001
|
| [9] | M. Martcheva, An Introduction to Mathematical Epidemiology, Springer, 2015. |
| [10] |
P. Van den Driessche, J. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180 (2002), 29–48. https://doi.org/10.1016/S0025-5564(02)00108-6 doi: 10.1016/S0025-5564(02)00108-6
|
| [11] |
D. Kiouach, Y. Sabbar, Stability and threshold of a stochastic SIRS epidemic model with vertical transmission and transfer from infectious to susceptible individuals, Discrete Dyn. Nat. Soc., 2018 (2018), 7570296. https://doi.org/10.1155/2018/7570296 doi: 10.1155/2018/7570296
|
| [12] |
D. Kiouach, Y. Sabbar, S. E. A. El-idrissi, New results on the asymptotic behavior of an SIS epidemiological model with quarantine strategy, stochastic transmission, and Levy disturbance, Math. Methods Appl. Sci., 44 (2021), 13468–13492. https://doi.org/10.1002/mma.7638 doi: 10.1002/mma.7638
|
| [13] |
Y. Sabbar, D. Kiouach, New method to obtain the acute sill of an ecological model with complex polynomial perturbation, Math. Methods Appl. Sci., 2022 (2022), forthcoming. https://doi.org/10.1002/mma.8654 doi: 10.1002/mma.8654
|
| [14] |
Y. Sabbar, A. Zeb, D. Kiouach, N. Gul, T. Sitthiwirattham, D. Baleanu et al., Dynamical bifurcation of a sewage treatment model with general higher-order perturbation, Results Phys., 39 (2022), 105799. https://doi.org/10.1016/j.rinp.2022.105799 doi: 10.1016/j.rinp.2022.105799
|
| [15] |
Y. Sabbar, A. Din, D. Kiouach, Predicting potential scenarios for wastewater treatment under unstable physical and chemical laboratory conditions: A mathematical study, Results Phys., 39 (2022), 105717. https://doi.org/10.1016/j.rinp.2022.105717 doi: 10.1016/j.rinp.2022.105717
|
| [16] |
Y. Sabbar, A. Khan, A. Din, Probabilistic analysis of a marine ecological system with intense variability, Mathematics, 10 (2022), 2262. https://doi.org/10.3390/math10132262 doi: 10.3390/math10132262
|
| [17] | D. Kiouach, Y. Sabbar, Developing new techniques for obtaining the threshold of a stochastic SIR epidemic model with 3-dimensional Lévy process, preprint, arXiv: 2002.09022. |
| [18] |
D. Kiouach, Y. Sabbar, The long-time behaviour of a stochastic SIR epidemic model with distributed delay and multidimensional Lévy jumps, Int. J. Biomath., 15 (2022), 2250004. https://doi.org/10.1142/S1793524522500048 doi: 10.1142/S1793524522500048
|
| [19] |
D. Kiouach, Y. Sabbar, Dynamic characterization of a stochastic SIR infectious disease model with dual perturbation, Int. J. Biomath., 14 (2021), 2150016. https://doi.org/10.1142/S1793524521500169 doi: 10.1142/S1793524521500169
|
| [20] |
D. Kiouach, Y. Sabbar, Ergodic stationary distribution of a stochastic hepatitis B epidemic model with interval-valued parameters and compensated poisson process, Comput. Math. Methods Med., 2020 (2020), 9676501. https://doi.org/10.1155/2020/9676501 doi: 10.1155/2020/9676501
|
| [21] |
A. Din, A. Khan, D. Baleanu, Stationary distribution and extinction of stochastic coronavirus (COVID-19) epidemic model, Chaos Solitons Fractals, 139 (2020), 110036. https://doi.org/10.1016/j.chaos.2020.110036 doi: 10.1016/j.chaos.2020.110036
|
| [22] |
A. Din, Y. Li, T. Khan, G. Zaman, Mathematical analysis of spread and control of the novel corona virus (COVID-19) in China, Chaos Solitons Fractals, 141 (2020), 110286. https://doi.org/10.1016/j.chaos.2020.110286 doi: 10.1016/j.chaos.2020.110286
|
| [23] |
A. Din and Y. Li and A. Yusuf, Delayed hepatitis B epidemic model with stochastic analysis, Chaos Solitons Fractals, 146 (2020), 110839. https://doi.org/10.1016/j.chaos.2021.110839 doi: 10.1016/j.chaos.2021.110839
|
| [24] |
D. Kiouach, Y. Sabbar, The threshold of a stochastic siqr epidemic model with Lévy jumps, Trends Biomath. Math. Model. Health Harvesting Popul. Dyn., 2019 (2019), 87–105. https://doi.org/10.1007/978-3-030-23433-1_7 doi: 10.1007/978-3-030-23433-1_7
|
| [25] |
D. Zhao, S. Yuan, Sharp conditions for the existence of a stationary distribution in one classical stochastic chemostat, Appl. Math. Comput., 339 (2018), 199–205. https://doi.org/10.1016/j.amc.2018.07.020 doi: 10.1016/j.amc.2018.07.020
|
| [26] |
N. Privault, L. Wang, Stochastic SIR Lévy jump model with heavy tailed increments, J. Nonlinear Sci., 31 (2021), 1–28. https://doi.org/10.1007/s00332-020-09670-5 doi: 10.1007/s00332-020-09670-5
|
| [27] | D. Kiouach, Y. Sabbar, Threshold analysis of the stochastic hepatitis b epidemic model with successful vaccination and Lévy jumps, in 2019 4th World Conference on Complex Systems (WCCS), IEEE, (2019). https://doi.org/10.1109/ICoCS.2019.8930709 |
| [28] |
Y. Sabbar, D. Kiouach, S. Rajasekar, S. E. A. El-idrissi, The influence of quadratic Lévy noise on the dynamic of an SIC contagious illness model: New framework, critical comparison and an application to COVID-19 (SARS-CoV-2) case, Chaos Solitons Fractals, 2022 (2022), 112110. https://doi.org/10.1016/j.chaos.2022.112110 doi: 10.1016/j.chaos.2022.112110
|
| [29] |
Y. Zhou, W. Zhang, Threshold of a stochastic SIR epidemic model with Levy jumps, Phys. A, 446 (2016), 204–216. https://doi.org/10.1016/j.physa.2015.11.023 doi: 10.1016/j.physa.2015.11.023
|
| [30] |
D. Zhao, S. Yuan, H. Liu, Stochastic dynamics of the delayed chemostat with Levy noises, Int. J. Biomath., 12 (2019), 1950056. https://doi.org/10.1142/S1793524519500566 doi: 10.1142/S1793524519500566
|



Figures(4) / Tables(1)

Asad Khan, Yassine Sabbar, Anwarud Din. Stochastic modeling of the Monkeypox 2022 epidemic with cross-infection hypothesis in a highly disturbed environment[J]. Mathematical Biosciences and Engineering, 2022, 19(12): 13560-13581. doi: 10.3934/mbe.2022633







 DownLoad:
DownLoad: