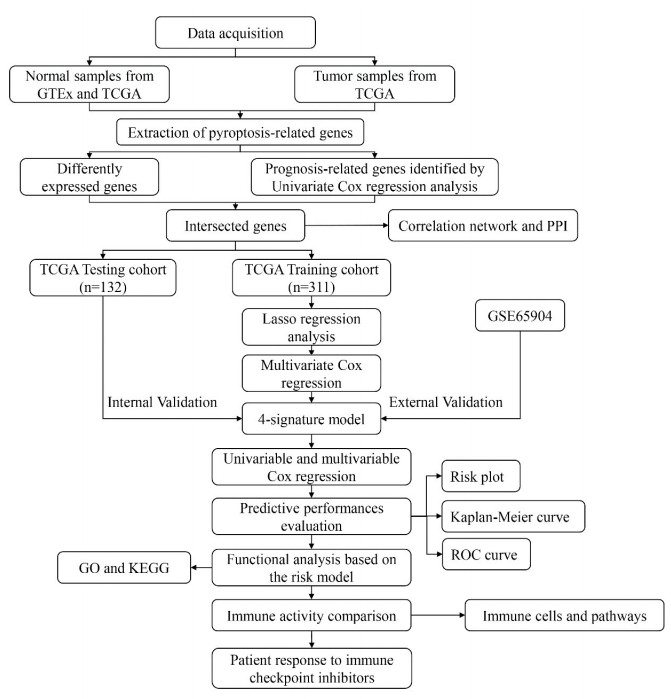
Skin cutaneous melanoma (SKCM) is one of the most malignant skin cancers and remains a health concern worldwide. Pyroptosis is a newly recognized form of programmed cell death and plays a vital role in cancer progression. We aim to construct a prognostic model for SKCM patients based on pyroptosis-related genes (PRGs). SKCM patients from The Cancer Genome Atlas (TCGA) were divided into training and validation cohorts. We used GSE65904 downloaded from GEO database as an external validation cohort. We performed Cox regression and the least absolute shrinkage and selection operator (LASSO) regression to identify prognostic genes and built a risk score. Patients were divided into high- and low-risk groups based on the risk score. Differently expressed genes (DEGs), immune cell infiltration and immune-related pathways activation were compared between the two groups. We established a model containing 4 PRGs, i.e., GSDMA, GSDMC, AIM2 and NOD2. The overall survival (OS) time was significantly different between the 2 groups. The risk score was an independent predictor for prognosis in both the uni- and multi-variable Cox regressions. Gene ontology (GO) and Kyoto Encylopedia of Genes and Genomes (KEGG) analyses showed that DEGs were enriched in immune-related pathways. Most types of immune cells were highly expressed in the low risk group. All immune pathways were significantly up-regulated in the low-risk group. In addition, low-risk patients had a better response to immune checkpoint inhibitors. Our novel pyroptosis-related gene signature could predict the prognosis of SKCM patients and their response to immune checkpoint inhibitors.
Citation: Zehao Niu, Yujian Xu, Yan Li, Youbai Chen, Yan Han. Construction and validation of a novel pyroptosis-related signature to predict prognosis in patients with cutaneous melanoma[J]. Mathematical Biosciences and Engineering, 2022, 19(1): 688-706. doi: 10.3934/mbe.2022031
Skin cutaneous melanoma (SKCM) is one of the most malignant skin cancers and remains a health concern worldwide. Pyroptosis is a newly recognized form of programmed cell death and plays a vital role in cancer progression. We aim to construct a prognostic model for SKCM patients based on pyroptosis-related genes (PRGs). SKCM patients from The Cancer Genome Atlas (TCGA) were divided into training and validation cohorts. We used GSE65904 downloaded from GEO database as an external validation cohort. We performed Cox regression and the least absolute shrinkage and selection operator (LASSO) regression to identify prognostic genes and built a risk score. Patients were divided into high- and low-risk groups based on the risk score. Differently expressed genes (DEGs), immune cell infiltration and immune-related pathways activation were compared between the two groups. We established a model containing 4 PRGs, i.e., GSDMA, GSDMC, AIM2 and NOD2. The overall survival (OS) time was significantly different between the 2 groups. The risk score was an independent predictor for prognosis in both the uni- and multi-variable Cox regressions. Gene ontology (GO) and Kyoto Encylopedia of Genes and Genomes (KEGG) analyses showed that DEGs were enriched in immune-related pathways. Most types of immune cells were highly expressed in the low risk group. All immune pathways were significantly up-regulated in the low-risk group. In addition, low-risk patients had a better response to immune checkpoint inhibitors. Our novel pyroptosis-related gene signature could predict the prognosis of SKCM patients and their response to immune checkpoint inhibitors.

| [1] |
R. L. Siegel, K. D. Miller, H. E. Fuchs, A. Jemal, Cancer statistics, CA Cancer J. Clin., 71 (2021), 7−33. doi: 10.3322/caac.20073. doi: 10.3322/caac.20073

|
| [2] |
K. D. Miller, L. Nogueira, A. B. Mariotto, J. H. Rowland, K. R. Yabroff, C. M. Alfano, et al., Cancer treatment and survivorship statistics, CA Cancer J. Clin., 69 (2019), 363−385. doi: 10.3322/caac.21235. doi: 10.3322/caac.21235
|
| [3] |
R. R. Kudchadkar, M. C. Lowe, M. K. Khan, S. M. McBrien, Metastatic melanoma, CA Cancer J. Clin., 70 (2020), 78−85. doi: 10.3322/caac.21599. doi: 10.3322/caac.21599
|
| [4] | M. Rastrelli, S. Tropea, C. R. Rossi, M. Alaibac, Melanoma: epidemiology, risk factors, pathogenesis, diagnosis and classification, In Vivo, 28 (2014), 1005−1011. |
| [5] |
X. Xia, X. Wang, Z. Cheng, W. Qin, L. Lei, J. Jiang, et al., The role of pyroptosis in cancer: pro-cancer or pro-"host"?, Cell Death Dis., 10 (2019), 650. doi: 10.1038/s41419-019-1883-8. doi: 10.1038/s41419-019-1883-8
|
| [6] |
P. Yu, H. Y. Wang, M. Tian, A. X. Li, X. S. Chen, X. L. Wang, et al., Eukaryotic elongation factor-2 kinase regulates the cross-talk between autophagy and pyroptosis in doxorubicin-treated human melanoma cells in vitro, Acta Pharmacol. Sin., 40 (2019), 1237−1244. doi: 10.1038/s41401-019-0222-z. doi: 10.1038/s41401-019-0222-z
|
| [7] |
S. Feng, D. Fox, S. M. Man, Mechanisms of gasdermin family members in inflammasome signaling and cell death, J. Mol. Biol., 430 (2018), 3068−3080. doi: 10.1016/j.jmb.2018.07.002. doi: 10.1016/j.jmb.2018.07.002
|
| [8] |
C. Lee, H. T. T. Do, J. Her, Y. Kim, D. Seo, I. Rhee, Inflammasome as a promising therapeutic target for cancer, Life Sci., 231 (2019), 116593. doi: 10.1016/j.lfs.2019.116593. doi: 10.1016/j.lfs.2019.116593
|
| [9] |
W. Hong, Y. Gu, R. Guan, D. Xie, H. Zhou, M. Yu, Pan-cancer analysis of the CASP gene family in relation to survival, tumor-infiltrating immune cells and therapeutic targets, Genomics, 112 (2020), 4304−4315. doi: 10.1016/j.ygeno.2020.07.026. doi: 10.1016/j.ygeno.2020.07.026
|
| [10] |
Q. Wei, K. Mu, T. Li, Y. Zhang, Z. Yang, X. Jia, et al., Deregulation of the NLRP3 inflammasome in hepatic parenchymal cells during liver cancer progression, Lab Invest., 94 (2014), 52−62. doi: 10.1038/labinvest.2013.126. doi: 10.1038/labinvest.2013.126
|
| [11] |
M. Hergueta-Redondo, D. Sarrió, Á Molina-Crespo, D. Megias, A. Mota, A. Rojo-Sebastian, et al., Gasdermin-B promotes invasion and metastasis in breast cancer cells, PLoS One, 9 (2014), e90099. doi: 10.1371/journal.pone.0090099. doi: 10.1371/journal.pone.0090099
|
| [12] |
W. J. Wang, D. Chen, M. Z. Jiang, B. Xu, X. W. Li, Y. Chu, et al., Downregulation of gasdermin D promotes gastric cancer proliferation by regulating cell cycle-related proteins, J. Dig. Dis., 19 (2018), 74−83. doi: 10.1111/1751-2980.12576. doi: 10.1111/1751-2980.12576
|
| [13] |
Y. Ye, Q. J. Dai, H. B. Qi, A novel defined pyroptosis-related gene signature for predicting the prognosis of ovarian cancer, Cell Death Discov., 7 (2021), 71. doi: 10.1038/s41420-021-00451-x. doi: 10.1038/s41420-021-00451-x
|
| [14] |
D. A. Erkes, W. Cai, I. M. Sanchez, T. J. Purwin, C. Rogers, C. O. Field, et al., Mutant BRAF and MEK inhibitors regulate the tumor immune microenvironment via pyroptosis, Cancer Discov., 10 (2020), 254−269. doi: 10.1158/2159-8290.CD-19-0672. doi: 10.1158/2159-8290.CD-19-0672
|
| [15] |
B. Zhou, J. Y. Zhang, X. S. Liu, H. Z. Chen, Y. L. Ai, K. Cheng, et al., Tom20 senses iron-activated ROS signaling to promote melanoma cell pyroptosis, Cell Res., 28 (2018), 1171−1185. doi: 10.1038/s41422-018-0090-y. doi: 10.1038/s41422-018-0090-y
|
| [16] |
J. Long, L. Zhang, X. Wan, J. Lin, Y. Bai, W. Xu, et al., A four-gene-based prognostic model predicts overall survival in patients with hepatocellular carcinoma, J. Cell Mol. Med., 22 (2018), 5928−5938. doi: 10.1111/jcmm.13863. doi: 10.1111/jcmm.13863
|
| [17] |
Y. Jia, Y. Chen, J. Liu, Prognosis-predictive signature and nomogram based on autophagy-related long non-coding RNAs for hepatocellular carcinoma, Front. Genet., 11 (2020), 608668. doi: 10.3389/fgene.2020.608668. doi: 10.3389/fgene.2020.608668
|
| [18] |
P. Charoentong, F. Finotello, M. Angelova, C. Mayer, M. Efremova, D. Rieder, et al., Pan-cancer immunogenomic analyses reveal genotype-immunophenotype relationships and predictors of response to checkpoint blockade, Cell Rep., 18 (2017), 248−262. doi: 10.1016/j.celrep.2016.12.019. doi: 10.1016/j.celrep.2016.12.019
|
| [19] |
D. Yan, S. Hu, Z. Zhou, S. Zeenat, F. Cheng, Y. Li, et al., Different chemical groups modification on the surface of chitosan nonwoven dressing and the hemostatic properties, Int. J. Biol. Macromol., 107 (2018), 463−469. doi: 10.1016/j.ijbiomac.2017.09.008. doi: 10.1016/j.ijbiomac.2017.09.008
|
| [20] |
C. Ci, B. Tang, D. Lyu, W. Liu, D. Qiang, X. Ji, et al., Overexpression of CDCA8 promotes the malignant progression of cutaneous melanoma and leads to poor prognosis, Int. J. Mol. Med., 43 (2019), 404−412. doi: 10.3892/ijmm.2018.3985. doi: 10.3892/ijmm.2018.3985
|
| [21] |
X. Zhang, X. Wen, N. Feng, A. Chen, S. Yao, X. Ding, et al., Increased expression of T-Box transcription factor protein 21 (TBX21) in skin cutaneous melanoma predicts better prognosis: A study based on The Cancer Genome Atlas (TCGA) and Genotype-Tissue Expression (GTEx) databases, Med. Sci. Monit., 26 (2020), e923087. doi: 10.12659/MSM.923087. doi: 10.12659/MSM.923087
|
| [22] |
B. Huang, W. Han, Z. F. Sheng, G. L. Shen, Identification of immune-related biomarkers associated with tumorigenesis and prognosis in cutaneous melanoma patients, Cancer Cell Int., 20 (2020), 195. doi: 10.21203/rs.2.23305/v3. doi: 10.21203/rs.2.23305/v3
|
| [23] |
Q. Wan, C. Liu, C. Liu, W. Liu, X. Wang, Z. Wang, Discovery and validation of a metastasis-related prognostic and diagnostic biomarker for melanoma based on single cell and gene expression datasets, Front. Oncol., 10 (2020), 585980. doi: 10.3389/fonc.2020.585980. doi: 10.3389/fonc.2020.585980
|
| [24] |
Y. Fang, S. Tian, Y. Pan, W. Li, Q. Wang, Y. Tang, et al., Pyroptosis: A new frontier in cancer, Biomed. Pharmacother., 121 (2020), 109595. doi: 10.1016/j.biopha.2019.109595. doi: 10.1016/j.biopha.2019.109595
|
| [25] |
A. A. Emran, H. Y. Tseng, M. C. Coleman, J. Tiffen, S. Cook, H. M. McGuire, et al., Do innate killing mechanisms activated by inflammasomes have a role in treating melanoma?, Pigment Cell Melanoma Res., 33 (2020), 660−670. doi: 10.1111/pcmr.12870. doi: 10.1111/pcmr.12870
|
| [26] |
H. Guo, J. B. Callaway, J. P. Ting, Inflammasomes: mechanism of action, role in disease, and therapeutics, Nat. Med., 21 (2015), 677−687. doi: 10.1038/nm.3893. doi: 10.1038/nm.3893
|
| [27] |
C. Rogers, D. A. Erkes, A. Nardone, A. E. Aplin, T. Fernandes-Alnemri, E. S. Alnemri, Gasdermin pores permeabilize mitochondria to augment caspase-3 activation during apoptosis and inflammasome activation, Nat. Commun., 10 (2019), 1689. doi: 10.1038/s41467-019-09397-2. doi: 10.1038/s41467-019-09397-2
|
| [28] |
M. Segovia, S. Russo, M. Jeldres, Y. D. Mahmoud, V. Perez, M. Duhalde, et al., Targeting TMEM176B enhances antitumor immunity and augments the efficacy of immune checkpoint blockers by unleashing inflammasome activation, Cancer Cell, 35 (2019), 767−781. e766. doi: 10.1016/j.ccell.2019.04.003. doi: 10.1016/j.ccell.2019.04.003
|
| [29] |
F. Ahmed, H. Y. Tseng, A. Ahn, D. Gunatilake, S. Alavi, M. Eccles, et al., Repurposing melanoma chemotherapy to activate inflammasomes in treatment of BRAF/MAPK inhibitor resistant melanoma, J. Invest. Dermatol., 2021 (2021). doi: 10.1016/j.jid.2021.09.030. doi: 10.1016/j.jid.2021.09.030
|
| [30] |
Y. Tan, Q. Chen, X. Li, Z. Zeng, W. Xiong, G. Li, et al., Correction to: Pyroptosis: a new paradigm of cell death for fighting against cancer, J. Exp. Clin. Cancer Res., 40 (2021), 219. doi: 10.1186/s13046-021-01959-x. doi: 10.1186/s13046-021-01959-x
|
| [31] |
J. Yu, Q. Wang, X. Zhang, Z. Guo, X. Cui, Mechanisms of neoantigen-targeted induction of pyroptosis and ferroptosis: From basic research to clinical applications, Front. Oncol., 11 (2021), 685377. doi: 10.3389/fonc.2021.685377. doi: 10.3389/fonc.2021.685377
|
| [32] |
L. M. Coussens, Z. Werb, Inflammation and cancer, Nature, 420 (2002), 860−867. doi: 10.1186/s12199-018-0740-1. doi: 10.1186/s12199-018-0740-1
|
| [33] |
K. Nagarajan, K. Soundarapandian, R. F. Thorne, D. Li, D. Li, Activation of pyroptotic cell death pathways in cancer: An alternative therapeutic approach, Transl. Oncol., 12 (2019), 925−931. doi: 10.1016/j.tranon.2019.04.010. doi: 10.1016/j.tranon.2019.04.010
|
| [34] |
C. Berkel, E. Cacan, Differential expression and Copy Number Variation of Gasdermin (GSDM) family members, pore-forming proteins in pyroptosis, in normal and malignant serous ovarian tissue, Inflammation, 2021 (2021). doi: 10.1007/s10753-021-01493-0. doi: 10.1007/s10753-021-01493-0
|
| [35] |
J. Ding, K. Wang, W. Liu, Y. She, Q. Sun, J. Shi, et al., Pore-forming activity and structural autoinhibition of the gasdermin family, Nature, 535 (2016), 111−116. doi: 10.1038/nature18590. doi: 10.1038/nature18590
|
| [36] |
J. Ruan, S. Xia, X. Liu, J. Lieberman, H. Wu, Cryo-EM structure of the gasdermin A3 membrane pore, Nature, 557 (2018), 62−67. doi: 10.1038/s41586-018-0058-6. doi: 10.1038/s41586-018-0058-6
|
| [37] |
N. Saeki, Y. Kuwahara, H. Sasaki, H. Satoh, T. Shiroishi, Gasdermin (Gsdm) localizing to mouse Chromosome 11 is predominantly expressed in upper gastrointestinal tract but significantly suppressed in human gastric cancer cells, Mamm. Genome., 11 (2000), 718−724. doi: 10.1007/s003350010138. doi: 10.1007/s003350010138
|
| [38] |
N. Saeki, T. Usui, K. Aoyagi, D. H. Kim, M. Sato, T. Mabuchi, et al., Distinctive expression and function of four GSDM family genes (GSDMA-D) in normal and malignant upper gastrointestinal epithelium, Genes Chromosomes Cancer, 48 (2009), 261−271. doi: 10.1002/gcc.20636. doi: 10.1002/gcc.20636
|
| [39] |
K. Watabe, A. Ito, H. Asada, Y. Endo, T. Kobayashi, K. Nakamoto, et al., Structure, expression and chromosome mapping of MLZE, a novel gene which is preferentially expressed in metastatic melanoma cells, Jpn. J. Cancer Res., 92 (2001), 140−151. doi: 10.1111/j.1349-7006.2001.tb01076.x. doi: 10.1111/j.1349-7006.2001.tb01076.x
|
| [40] |
J. Hou, R. Zhao, W. Xia, C. W. Chang, Y. You, J. M. Hsu, et al., PD-L1-mediated gasdermin C expression switches apoptosis to pyroptosis in cancer cells and facilitates tumour necrosis, Nat. Cell Biol., 22 (2020), 1264−1275. doi: 10.1038/s41556-020-0575-z. doi: 10.1038/s41556-020-0575-z
|
| [41] |
M. Miguchi, T. Hinoi, M. Shimomura, T. Adachi, Y. Saito, H. Niitsu, et al., Gasdermin C Is upregulated by inactivation of transforming growth factor β receptor Type Ⅱ in the presence of mutated apc, promoting colorectal cancer proliferation, PLoS One, 11 (2016), e0166422. doi: 10.1371/journal.pone.0166422. doi: 10.1371/journal.pone.0166422
|
| [42] |
J. Wei, Z. Xu, X. Chen, X. Wang, S. Zeng, L. Qian, et al., Overexpression of GSDMC is a prognostic factor for predicting a poor outcome in lung adenocarcinoma, Mol. Med. Rep., 21 (2020), 360−370. doi: 10.3892/mmr.2019.10837. doi: 10.3892/mmr.2019.10837
|
| [43] |
A. Ito, K. Watabe, Y. Koma, Y. Kitamura, An attempt to isolate genes responsible for spontaneous and experimental metastasis in the mouse model, Histol. Histopathol., 17 (2002), 951−959. doi: 10.14670/HH-17.951. doi: 10.14670/HH-17.951
|
| [44] |
B. R. Sharma, R. Karki, T. D. Kanneganti, Role of AIM2 inflammasome in inflammatory diseases, cancer and infection, Eur. J. Immunol., 49 (2019), 1998−2011. doi: 10.1002/eji.201848070. doi: 10.1002/eji.201848070
|
| [45] |
M. Xu, J. Wang, H. Li, Z. Zhang, Z. Cheng, AIM2 inhibits colorectal cancer cell proliferation and migration through suppression of Gli1, Aging (Albany NY), 13 (2020), 1017−1031. doi: 10.18632/aging.202226. doi: 10.18632/aging.202226
|
| [46] |
M. Farshchian, L. Nissinen, E. Siljamäki, P. Riihilä, M. Piipponen, A. Kivisaari, et al., Tumor cell-specific AIM2 regulates growth and invasion of cutaneous squamous cell carcinoma, Oncotarget, 8 (2017), 45825−45836. doi: 10.18632/oncotarget.17573. doi: 10.18632/oncotarget.17573
|
| [47] |
R. Caruso, N. Warner, N. Inohara, G. Núñez, NOD1 and NOD2: signaling, host defense, and inflammatory disease, Immunity, 41 (2014), 898−908. doi: 10.1016/j.immuni.2014.12.010. doi: 10.1016/j.immuni.2014.12.010
|
| [48] |
X. Ma, Y. Qiu, Y. Sun, L. Zhu, Y. Zhao, T. Li, et al., NOD2 inhibits tumorigenesis and increases chemosensitivity of hepatocellular carcinoma by targeting AMPK pathway, Cell Death Dis., 11 (2020), 174. doi: 10.1038/s41419-020-2368-5. doi: 10.1038/s41419-020-2368-5
|
| [49] |
S. A. Gurses, S. Banskar, C. Stewart, B. Trimoski, R. Dziarski, D. Gupta, Nod2 protects mice from inflammation and obesity-dependent liver cancer, Sci. Rep., 10 (2020), 20519. doi: 10.1038/s41598-020-77463-7. doi: 10.1038/s41598-020-77463-7
|
| [50] |
Y. Zhou, L. Hu, W. Tang, D. Li, L. Ma, H. Liu, et al., Hepatic NOD2 promotes hepatocarcinogenesis via a RIP2-mediated proinflammatory response and a novel nuclear autophagy-mediated DNA damage mechanism, J. Hematol. Oncol., 14 (2021), 9. doi: 10.1186/s13045-020-01028-4. doi: 10.1186/s13045-020-01028-4
|
| [51] |
T. Fujimura, K. Yamasaki, T. Hidaka, Y. Ito, S. Aiba, A synthetic NOD2 agonist, muramyl dipeptide (MDP)-Lys (L18) and IFN-β synergistically induce dendritic cell maturation with augmented IL-12 production and suppress melanoma growth, J. Dermatol. Sci., 62 (2011), 107−115. doi: 10.1016/j.jdermsci.2011.02.002. doi: 10.1016/j.jdermsci.2011.02.002
|
| [52] |
A. Ladányi, Prognostic and predictive significance of immune cells infiltrating cutaneous melanoma, Pigment Cell Melanoma Res., 28 (2015), 490−500. doi: 10.1111/pcmr.12371. doi: 10.1111/pcmr.12371
|
| [53] |
S. R. Selitsky, L. E. Mose, C. C. Smith, S. Chai, K. A. Hoadley, D. P. Dittmer, et al., Prognostic value of B cells in cutaneous melanoma, Genome Med., 11 (2019), 36. doi: 10.1186/s13073-019-0647-5. doi: 10.1186/s13073-019-0647-5
|
| [54] |
M. Tucci, A. Passarelli, F. Mannavola, C. Felici, L. S. Stucci, M. Cives, et al., Immune system evasion as hallmark of melanoma progression: The role of dendritic cells, Front. Oncol., 9 (2019), 1148. doi: 10.3389/fonc.2019.01148. doi: 10.3389/fonc.2019.01148
|
| [55] |
Z. Zhang, Y. Zhang, S. Xia, Q. Kong, S. Li, X. Liu, et al., Gasdermin E suppresses tumour growth by activating anti-tumour immunity, Nature, 579 (2020), 415−420. doi: 10.1038/s41586-020-2071-9. doi: 10.1038/s41586-020-2071-9
|
| [56] |
Q. Wang, Y. Wang, J. Ding, C. Wang, X. Zhou, W. Gao, et al., A bioorthogonal system reveals antitumour immune function of pyroptosis, Nature, 579 (2020), 421−426. doi: 10.1038/s41586-020-2079-1. doi: 10.1038/s41586-020-2079-1
|
| [57] |
S. L. Topalian, J. M. Taube, R. A. Anders, D. M. Pardoll, Mechanism-driven biomarkers to guide immune checkpoint blockade in cancer therapy, Nat. Rev. Cancer, 16 (2016), 275−287. doi: 10.1038/nrc.2016.36. doi: 10.1038/nrc.2016.36
|
| [58] |
C. C. Zhang, C. G. Li, Y. F. Wang, L. H. Xu, X. H. He, Q. Z. Zeng, et al., Chemotherapeutic paclitaxel and cisplatin differentially induce pyroptosis in A549 lung cancer cells via caspase−3/GSDME activation, Apoptosis, 24 (2019), 312−325. doi: 10.1007/s10495-019-01515-1. doi: 10.1007/s10495-019-01515-1
|
| [59] |
E. Yue, G. Tuguzbaeva, X. Chen, Y. Qin, A. Li, X. Sun, et al., Anthocyanin is involved in the activation of pyroptosis in oral squamous cell carcinoma, Phytomedicine, 56 (2019), 286−294. doi: 10.1016/j.phymed.2018.09.223. doi: 10.1016/j.phymed.2018.09.223
|
| [60] |
R. Ghiman, M. Nistor, M. Focșan, A. Pintea, S. Aștilean, D. Rugina, Fluorescent polyelectrolyte system to track anthocyanins delivery inside melanoma cells, Nanomaterials (Basel), 11 (2021), 782. doi: 10.3390/nano11030782. doi: 10.3390/nano11030782
|
| [61] |
A. Ashworth, ATR inhibitors and paclitaxel in melanoma, Clin. Cancer Res., 27 (2021), 4667−4668. doi: 10.1158/1078-0432.CCR-21-1778. doi: 10.1158/1078-0432.CCR-21-1778
|
| [62] |
J. J. Li, J. H. Wang, Y. Dingv, D. D. Li, X. Z. Wen, J. J. Zhao, et al., Efficacy and safety of anti-PD-1 inhibitor combined with nab-paclitaxel in Chinese patients with refractory melanoma, J. Cancer Res. Clin. Oncol., 2021 (2021). doi: 10.1007/s00432-021-03700-9. doi: 10.1007/s00432-021-03700-9
|
| [63] |
L. Ai, A. Xu, J. Xu, Roles of PD-1/PD-L1 pathway: Signaling, cancer, and beyond, Adv. Exp. Med. Biol., 1248 (2020), 33−59. doi: 10.1007/978-981-15-3266-5_3. doi: 10.1007/978-981-15-3266-5_3
|
| [64] |
S. Kleffel, C. Posch, S. R. Barthel, H. Mueller, C. Schlapbach, E. Guenova, et al., Melanoma cell-intrinsic PD-1 receptor functions promote tumor growth, Cell, 162 (2015), 1242−1256. doi: 10.1016/j.cell.2015.08.052. doi: 10.1016/j.cell.2015.08.052
|
| [65] |
B. Rowshanravan, N. Halliday, D. M. Sansom, CTLA-4: a moving target in immunotherapy, Blood, 131 (2018), 58−67. doi: 10.1182/blood-2017-06-741033. doi: 10.1182/blood-2017-06-741033
|
| [66] |
F. S. Hodi, S. J. O'Day, D. F. McDermott, R. W. Weber, J. A. Sosman, J. B. Haanen, et al., Improved survival with ipilimumab in patients with metastatic melanoma, N. Engl. J. Med., 363 (2010), 711−723. doi: 10.1056/NEJMoa1003466. doi: 10.1056/NEJMoa1003466
|
| [67] |
T. N. Gide, J. S. Wilmott, R. A. Scolyer, G. V. Long, Primary and acquired resistance to immune checkpoint inhibitors in metastatic melanoma, Clin. Cancer Res., 24 (2018), 1260−1270. doi: 10.1016/j.jtho.2021.01.131. doi: 10.1016/j.jtho.2021.01.131
|






Figures(7)

Zehao Niu, Yujian Xu, Yan Li, Youbai Chen, Yan Han. Construction and validation of a novel pyroptosis-related signature to predict prognosis in patients with cutaneous melanoma[J]. Mathematical Biosciences and Engineering, 2022, 19(1): 688-706. doi: 10.3934/mbe.2022031







 DownLoad:
DownLoad: